|
 CICLO VITAL.
CICLO VITAL.
Los ascomicetos son hongos con micelio haploide que pueden reproducirse
tanto sexual como asexualmente. Las levaduras se reproducen asexualmente por
gemación, aunque a veces
sus células pueden quedar enlazadas, formando un
pseudomicelio. Como norma
general, el anamorfo sirve para propagarse a gran
velocidad en la época favorable, siempre que las condiciones ambientales sean
más o menos estables y uniformes (en un cultivo, por ejemplo). Cuando los buenos
tiempos se acaban, se lleva a cabo la reproducción sexual. En el caso de los
fitoparásitos, resulta corriente que el teleomorfo se forme en los restos de la
cosecha, sirva para pasar la época desfavorable y sea la fuente de inóculo para
la siguiente estación. En este inóculo habrá ocurrido la recombinación genética,
por lo que se incrementarán las posibilidades de adaptarse a los cambios (nuevas
variedades de plantas, nuevos fitosanitarios...). Muchos de estos hongos, sin
embargo, tienden a perder la fase sexual, reemplazándola por estrategias
parasexuales. En este caso, la época desfavorable es superada por el micelio
(latente, en hospedantes alternativos, etc.), los
conidios,
clamidósporas o
esclerocios. Los líquenes
tienen diversas estrategias de reproducción asexual que no trataremos aquí.
Fase sexual.
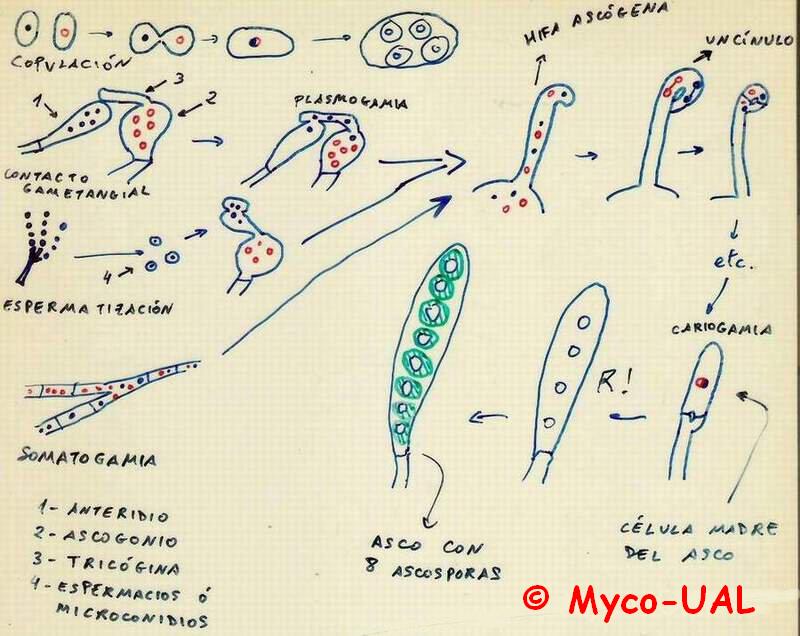
El primer paso para la reproducción sexual es la plasmogamia, o fusión de
células o estructuras reproductoras (fig. 1). En las levaduras, la plasmogamia
ocurre por copulación de gametos. En los ascomicetos más complejos se da por
medio de contacto entre un anteridio y un ascogonio, por espermatización (unos
espermacios llegan hasta el ascogonio o, mejor dicho, hasta una prolongación
de éste llamada tricógina), o bien por simple somatogamia. En cualquier caso,
la cariogamia (fusión de los núcleos para dar uno diploide) no ocurre
inmediatamente después de la plasmogamia, sino que se da lugar a unas hifas
dicarióticas (es decir, con dos tipos diferentes de núcleos), llamadas
hifas ascógenas. Éstas crecen mediante formación de uncínulos, hasta que en un momento
dado en una célula terminal ocurre la cariogamia, convirtiéndose en la
célula madre del asco. Seguidamente ocurre la meiosis, y los 4 núcleos resultantes se
convierten en 8 tras una mitosis. Cada núcleo se rodea de citoplasma y pared,
y da lugar a una ascóspora (hay especies que no presentan las 8 típicas
ascósporas; algunas pueden abortar o, por el contrario, dividirse aún más).
|
|
|
Figura 1:
Plasmogamia y cariogamia en ascomicetos (explicación en
el texto).
|
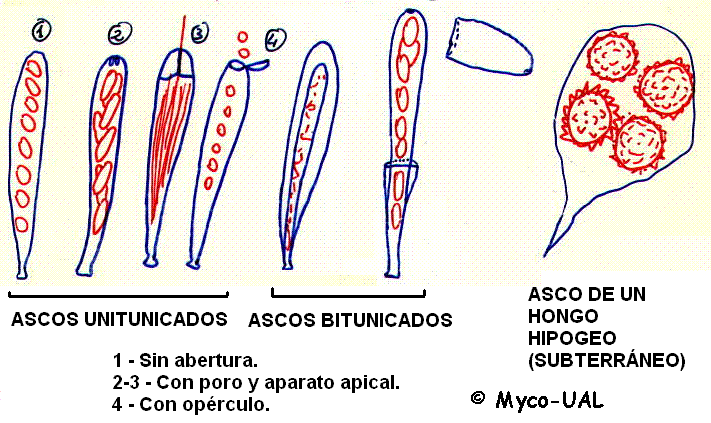
Los ascos se dividen en tres grandes grupos. Los
prototunicados tienen paredes
delicadas y liberan sus esporas por delicuescencia (es decir, por disolución de
la pared), para ser dispersadas de forma pasiva (por pequeños animales, en muchos
casos). En el resto de ascos, la pared consta de dos capas: exotúnica y
endotúnica. En los unitunicados,
ambas capas están firmemente adheridas, mientras que en los
bitunicados la endotúnica se hincha
y la exotúnica, más rígida, se rompe al madurar. En ambos casos las ascósporas se suelen
liberar de forma violenta. La mayor parte de ellas tienen forma de elipsoide a
elíptico-fusoide (y en muchas ocasiones recuerdan a la de un supositorio) para poder salir a
presión por la estrecha abertura del asco (que puede ser desde un vulgar desgarro
hasta un poro o un opérculo, tal como se muestra en la fig. 2). Los ascos de los
hongos hipogeos (subterráneos), como las trufas, no sin disparados violentamente,
ya que se dispersan tras ser ingeridos por mamíferos u otros animales, pasando por
el tubo digestivo y depositándose con los excrementos (Fig. 2).
|
|
|
Figura 2:
Tipos de ascos.
|
Los ascos pueden aparecer desnudos, como en las levaduras o en
Taphrina, pero lo más
normal es que se organicen en cuerpos fructíferos llamados ascocarpos o
ascomas. Los ascocarpos más corrientes son los
cleistotecios, esféricos y
cerrados; los peritecios,
desde redondeados hasta con forma de botella,
con un poro apical, el ostiolo, por donde se liberan las esporas; los
apotecios,
ascomas en forma de disco o plato, con o sin pie; y los
ascostromas o pseudotecios,
huecos que surgen en un estroma, dentro de los cuales se forman los ascos. Éstos
pueden aparecer ordenados en una capa fértil, denominada
himenio. Entre los ascos pueden
aparecer diversas hifas estériles; su conjunto recibe el nombre de hamatecio. Las
más conocidas son las paráfisis,
aunque hay otros tipos.
Ciclo parasexual.
La reproducción asexual, a causa de su economía y rapidez, es sumamente práctica
para la veloz dispersión de una especie en ambientes estables. Sin embargo, el sexo
resulta ventajoso en ambientes cambiantes, ya que proporciona variabilidad genética
por medio de la recombinación. Como ya se dijo, los hongos suelen aprovechar lo mejor
de ambas estrategias: multiplicación vegetativa en época de vacas gordas, y sexo
cuando las cosas se ponen feas. Sin embargo, muchos hongos imperfectos que aparentemente
no se propagan sexualmente sobreviven de un año para otro. Es más, son muy variables
(como el agricultor ha comprobado para su desgracia, es fácil la aparición de cepas
resistentes a fungicidas, o que ataquen a variedades de plantas supuestamente inatacables).
¿Cómo lo hacen? Una posibilidad es la del ciclo parasexual, que no requiere meiosis
ni la formación de gametangios especializados. Dicho ciclo fue propuesto en 1952 por
Pontecorvo y Roper para Aspergillus nidulans.
Veamos un breve resumen del ciclo parasexual:
- Para que haya parasexualidad se requiere primero la existencia de un
heterocarionte, es decir, un hongo con núcleos de distinto tipo en
el mismo micelio. Esto se puede lograr bien por mutación o, más frecuentemente,
porque dos micelios diferentes se anastomosan y uno le pasa sus núcleos
al otro. Debe quedar claro que cuando hablamos de núcleos "distintos" nos
referimos a que poseen alelos distintos para el mismo gen. Y no olvidemos
que se trata de nucleos haploides, con un único juego de cromosomas.
- Dentro de las células del heterocarionte, por azar (o sea, sin que
el hongo se tome la molestia de fabricar estructuras especiales para ello)
puede darse la cariogamia (fusión) entre núcleos distintos.
Obtendríamos así algunos núcleos diploides, es decir, con dos juegos de
cromosomas.
- Los núcleos diploides resultantes se dividen por mitosis, pero en ese
proceso pueden ocurrir entrecruzamientos mitóticos entre cromosomas
homólogos (y, por tanto, recombinación genética).
- Estos núcleos diploides pueden sufrir mitosis aberrantes. Así, a partir
de un núcleo con 2n cromosomas pueden obtenerse
núcleos aneuploides,
con 2n+1 y 2n-1 cromosomas. Pero estos aneuploides son
inestables...
- Poco a poco, a partir de los aneuploides pueden volver a formarse
núcleos haploides por pérdida de cromosomas. Estos nuevos núcleos haploides
tendrán una mezcla de caracteres respecto de los núcleos originales. Si van
a parar a un conidio, el individuo resultante tras la germinación tendrá
una dotación genética distinta a la parental. En suma, ha habido recombinación,
pero sin meiosis, sexo ni nada que se le parezca.
El proceso es 500 veces menos efectivo que el sexo, pero si se tiene en cuenta
que un individuo puede producir millones de conidios, resulta que funciona muy bien.
Se desconoce el papel real de la parasexualidad en la naturaleza;
hay autores que opinan que no es significativa, ya que estos hongos suelen presentar
mecanismos de incompatibilidad vegetativa que evitan la heterocariosis.
Estructuras reproductoras asexuales.
Los hongos anamorfos se reproducen asexualmente por gemación (en levaduras),
por simple fragmentación del micelio o por medio de conidios. Éstos son producidos
mediante fragmentación o gemación por unas células conidiógenas, que suelen aparecer
sobre unas hifas especializadas, los conidióforos (no en todos los casos esta
distinción queda clara; véase el género Alternaria). En otras ocasiones, estas
estructuras aparecen agrupadas en cuerpos más complejos, denominados conidiomas.
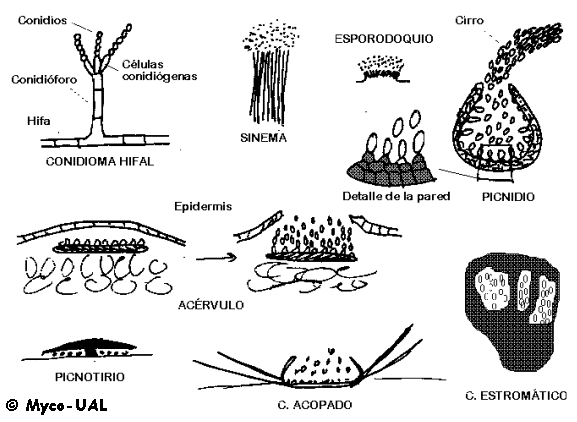
Sus principales tipos se ilustran en la fig. 3:
|
|
|
Figura 3:
Tipos de conidiomas.
|
- Conidiomas
hifales: Se trata de conidióforos aislados.
- Sinemas o
coremios: Conidióforos unidos a modo de penacho o cerdas
de un pincel.
- Esporodoquios:
Conidióforos cortos, unidos de forma más compacta que la anterior,
a veces dispuestos sobre un estroma y que adoptan un aspecto almohadillado.
- Picnidios:
Conidiomas cerrados, globosos o en forma de botella, abiertos por un
ostiolo y repletos de conidios. A menudo éstos salen en forma de
cirro.
- Acérvulos (Fig. 3): Conidiomas inmersos en el hospedante,
que consisten en un estroma plano, sobre el que se disponen conidióforos
cortos o células conidiógenas de forma similar a las velitas en un pastel
de cumpleaños. La presión de los conidios al crecer rompe la epidermis del
hospedante (por cierto, si un hongo de estos se hace crecer en
agar forma esporodoquios, ya que no hay epidermis que romper...).
- Conidiomas acopados (Fig. 3): Son superficiales, en forma de
cuenco, normalmente rodeados de pelos.
- Picnotirios (Fig. 3): Conidiomas en forma de escudo, con los
conidios debajo.
- Conidiomas estromáticos (Fig. 3): Los que por su forma no pueden
incluirse en los grupos anteriores.
La clasificación tradicional, puramente morfológica, se basaba en el tipo de conidiomas.
Dentro de la subdivisión Deuteromycotina se distinguían las siguientes clases:
- Clase Blastomycetes: levaduras asexuales (que en algunos casos pueden producir micelio).
- Clase Hyphomycetes: hongos con conidiomas hifales, sinemas o esporodoquios (también se
incluyen aquí los "micelios estériles", que ni siquiera se molestan en formar conidios).
- Clase Coelomycetes: hongos con otro tipo de conidiomas (picnidios, sobre todo).
Los órdenes se basaban en el tipo concreto de conidiomas. A niveles
taxonómicos más bajos se seguía el criterio de considerar la forma y
color de los conidios.
La clasificación anterior resultaba claramente artificial (aunque tan
práctica que todo el mundo la utilizaba). Por ello, algunos investigadores
buscaron otros criterios para ordenar coherentemente a estos hongos. Hughes
(1957) propuso que la taxonomía de Deuteromycotina habría de basarse en la
conidiogénesis, es decir, el proceso de formación de los conidios. Este
sistema, aparentemente más natural, fue mejorado por M.B. Ellis (1971) y
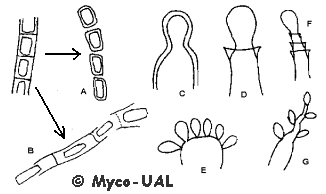
muchos otros. Aparece resumido en la fig. 4.
|
|
|
Figura 4:
Tipos de conidiogénesis (explicación en el texto).
|
Hay dos grandes tipos de conidiogénesis: tálica (fig. 4 A,B), por
fragmentación del micelio, y blástica (fig. 4 C,D), por gemación. En
ambos casos se añade el prefijo holo- (fig. 4 A,C) si todas las capas
de la pared de la célula conidiógena intervienen en la formación del
conidio, o entero- (fig. 4 B,D), si sólo interviene la parte interna de
la pared (las células conidiógenas enteroblásticas se llaman fiálides).
La célula conidiógena puede complicar aún más las cosas: a veces produce
los conidios en cadena;
en otras ocasiones los conidios se forman simultáneamente en varios lugares
a la vez (fig. 4 E; un buen ejemplo es
Botrytis). En
ciertos géneros, la célula conidiógena
prolifera (es decir,
crece tras formar un conidio, como en la fig. 4 F-G). En el caso de la fig.
4F, las proliferaciones siguen el mismo eje del crecimiento, y la célula
conidiógena se denomina anélide.
|
|
|
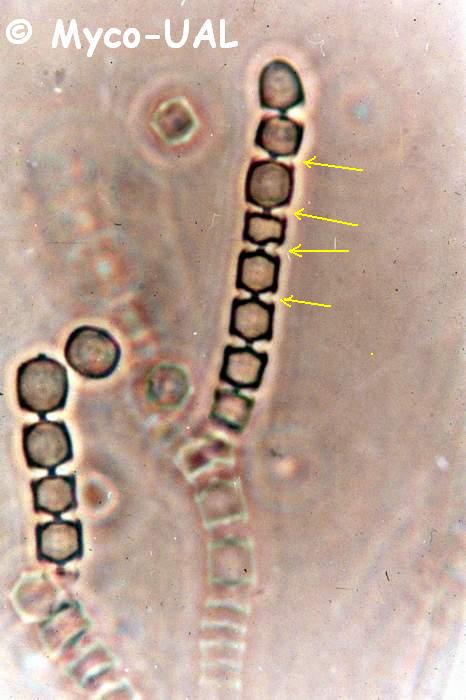
Figura 5:
Conidiogénesis enterotálica en Coremiella cubispora.
Véanse los restos de la parte externa de la pared entre
algunos conidios (flechas).
|
La conidiogénesis parecía un magnífico criterio para ordenar estos hongos,
y se publicaron numerosos libros y claves de acuerdo con ella. El único
problema es que se trata de un carácter poco fiable: en muchos hongos es
difícil de observar, puede cambiar según la edad del micelio, o la temperatura...
En resumen, dada la artificialidad de este grupo a veces es mejor emplear los
criterios clásicos, que resultan más fáciles de manejar. De todos modos, se
recomienda la consulta de la última edición del Dictionary, donde se definen e
ilustran detalles de conidiogénesis y modos de crecimiento de las paredes de
los conidios.
Volver a Fungi.
|