 Los orígenes de las células gliales en la literatura de neuroanatomía
se extiende desde el año de 1846, en el que aparecieron los trabajos
de Rudolph Virchow. Este hecho histórico ha sido documentado ampliamente
por Glees, Kuffler y Nicholls. La definición de células gliales
se deriva esencialmente de un proceso de exclusión. Pueden definirse
como células del tejido nervioso que no son neuronas. Sin embargo,
las células gliales poseen también uno o dos componentes
bioquímicos por los que pueden distinguirse de otras células
(Bradford, 1988).
Los orígenes de las células gliales en la literatura de neuroanatomía
se extiende desde el año de 1846, en el que aparecieron los trabajos
de Rudolph Virchow. Este hecho histórico ha sido documentado ampliamente
por Glees, Kuffler y Nicholls. La definición de células gliales
se deriva esencialmente de un proceso de exclusión. Pueden definirse
como células del tejido nervioso que no son neuronas. Sin embargo,
las células gliales poseen también uno o dos componentes
bioquímicos por los que pueden distinguirse de otras células
(Bradford, 1988).
Los matices funcionales
atribuidos en la actualidad a las células neurogliales son:
* Soporte mecánico
de las neuronas (astrocitos y oligodendrocitos).
* Producción
de la vaina de mielina (oligodendrocitos).
* Captación
rápida y, por tanto, inactivación de neurotransmisores químicos
liberados por las neuronas (astrocitos).
* Formación
del tejido cicatricial después de lesiones cerebrales (astrocitos).
* Eliminación
de residuos de tejido local después de la muerte celular (astrocitos)
(Gorovits et al., 1997; Newcomb et al., 1997).
* Constitución
de un sistema de fibras entre la sangre y las neuronas (astrocitos).
* Control de la composición
del líquido extracelular. Niveles de los iones de potasio y calcio
(astrocitos) (Bradford, 1988).
Tipos
de células gliales
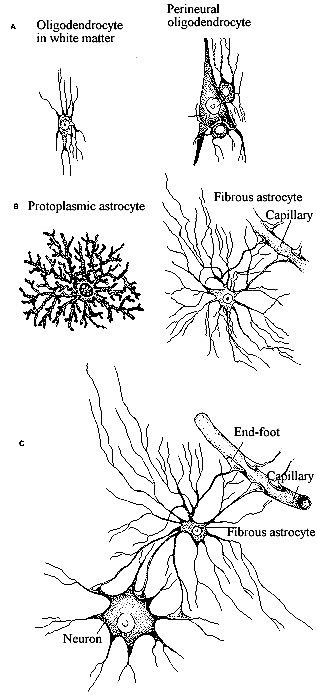 Teniendo en cuenta el tamaño, forma y situación se
distinguen varios subgrupos de células, gliales. La gran familia
de células gliales puede dividirse en macroglías, que
son las células de mayor tamaño, y microglías las
menores. Entre la macroglía está la glía radial, astroglía,
oligodendroglía y glía periférica (Bradford, 1988).
Teniendo en cuenta el tamaño, forma y situación se
distinguen varios subgrupos de células, gliales. La gran familia
de células gliales puede dividirse en macroglías, que
son las células de mayor tamaño, y microglías las
menores. Entre la macroglía está la glía radial, astroglía,
oligodendroglía y glía periférica (Bradford, 1988).
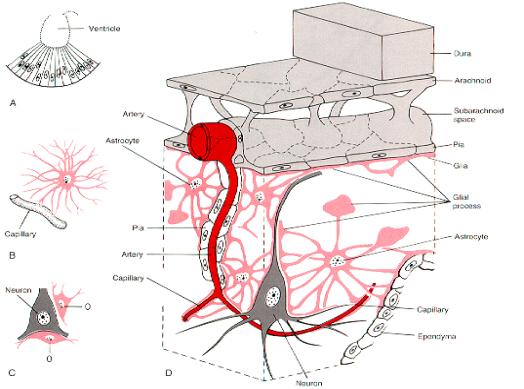
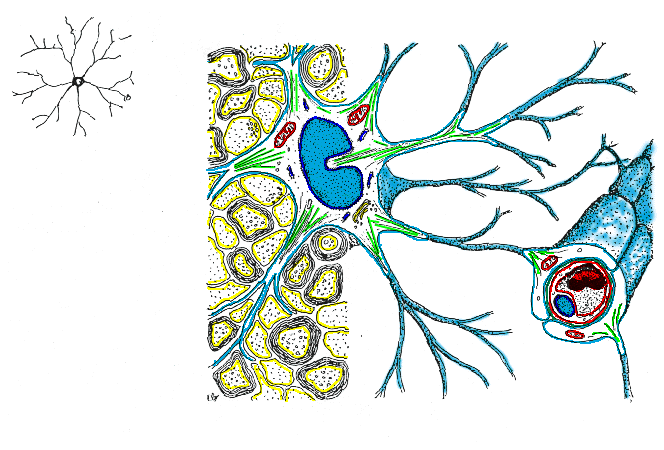
Las células
gliales de mayor tamaño son los astrocitos. Su pericarión,
de 18 a 20 mm
de diámetro, posee un núcleo pálido de gran tamaño,
gránulos de glucógeno y en algunos casos abundantes filamentos
gliales (gliofilamentos). Sus prolongaciones membranosas, que son largas
y estrechas, proporcionan la forma característica de estrella señalada
en su nombre. Estas estructuras delgadas y las dilataciones que se aprecian
en sus extremidades (pies terminales) rodean la superficie de los capilares
sanguíneos, dejando solamente unos huecos pequeños para la
difusión de las sustancias desde los endotelios capilares a través
de la lámina basal y los espacios extracelulares restringidos del
encéfalo. Los pies terminales, que son una característica
especial de los astrocitos se hallan también superpuestos a las
membranas neuronales y a las membranas piales que cubren la superficie
encefálica externa. La oligodendroglía constituye otro subgrupo
glial importante. Estas células se distinguen claramente de los
astrocitos por sus pequeños cuerpos celulares (3 a 5 mm
de diámetro) con pocas prolongaciones celulares. La oligodendroglía
se halla en la sustancia blanca y en la sustancia gris. La de la sustancia
blanca conocida como oligodendroglía interfascicular, es responsable
de la formación de la vaina de mielina aislante alrededor de los
axones; en la sustancia blanca del cerebro en desarrollo, sus
prolongaciones celulares se distinguen continuas con las vainas de mielina
que están produciendo ininterrumpidamente.
En la sustancia gris la oligodendroglía satélite se halla
frecuentemente en íntima relación con los cuerpos celulares.
La microglía es la neuroglía más pequeña, con
diámetro de 2 a 3 mm,
por lo que pueden distinguirse fácilmente de la macroglía.
Su variable morfología hace que sea difícil de definir y
estudiar, sin embargo, estas son las células encargadas del sistema
inmune del SNC (Bradford, 1988).
 Estas células se distinguen por la facilidad con que son teñidas
con las sales de plata, por sus núcleos pequeños y alargados,
por su escaso citoplasma y por su multitud de prolongaciones celulares
pequeñas (Bradford, 1988). Cuando estas células se exponen
a estímulos que provocan inflamación u otro tipo de insulto
al tejido nervioso, la microglía se transforma rápidamente
desde un estado de reposo pasivo a una fagocitosis activa acompañada
de proliferación y migración por el tejido. Es en esta fase
cuyo su similitud con los macrófagos es evidente (Bradford, 1988).
Estas células se distinguen por la facilidad con que son teñidas
con las sales de plata, por sus núcleos pequeños y alargados,
por su escaso citoplasma y por su multitud de prolongaciones celulares
pequeñas (Bradford, 1988). Cuando estas células se exponen
a estímulos que provocan inflamación u otro tipo de insulto
al tejido nervioso, la microglía se transforma rápidamente
desde un estado de reposo pasivo a una fagocitosis activa acompañada
de proliferación y migración por el tejido. Es en esta fase
cuyo su similitud con los macrófagos es evidente (Bradford, 1988).
Las células gliales radiales, descritas por primera vez a finales
del siglo pasado, han sido consideradas en la actualidad como un grupo
especial de neuroglía que tiene una importancia considerable.
Estas células aparecen durante el desarrollo encefálico para
transformarse después en otros tipos gliales (por ejemplo, astrocitos)
a medida que madura el cerebro. Muchas de estas células poseen una
forma alargada, frecuentemente bipolar, con dos o más prolongaciones
celulares importantes que se extienden a distancias relativamente largas
a través de tejido nervioso y que terminan frecuentemente en las
membranas superficiales o en las paredes de los vasos sanguíneos.
Al menos hay dos tipos de glía radial que sobreviven en el cerebro
adulto humano: las células de Müller y la glía
de Bergman. Las células de Müller se hallan en la retina y
muestran una morfología y disposición características;
son muy alargadas y se extienden entre las membranas limitantes interna
y externa de la retina. La glía de Bergman conformada por
células gliales peculiares, situadas en la capa molecular en la
corteza cerebelosa de los mamíferos proyecta varias prolongaciones
celulares alargadas a través de la capa molecular hasta la superficie
interna de la membrana pial, donde forman pies terminales cónicos.
Los cuerpos celulares de la glía de Bergman están localizados
en la capa de células de Purkinje (voluminosas neuronas ramificadas
de la capa media de la corteza cerebelar) (Bradford, 1988).
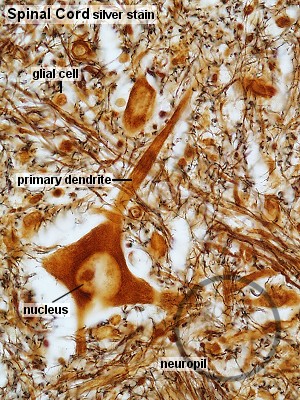
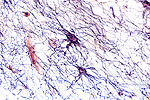 Los astrocitos como su nombre lo sugiere, son células en forma
de estrella, con numerosas prolongaciones citoplásmicas ramificadas.
Sus núcleos son grandes, ovoides o esféricos y tienen color
pálido, con pocos gránulos finos de cromatina distribuidos
principalmente en la periferia en unión íntima con la membrana
nuclear. Los nucléolos no son patentes. El citoplasma incluye los
organelos corrientes y puede observarse claramente un centrosoma. Puede
haber pigmento lipocrómico (Lesson, 1976). Sus prolongaciones celulares
fibrosas características, así como su pericarión,
están rellenas de gliofilamentos cuyo componente proteico más
importante, llamado proteína ácida fibrilar de la glía
(PAFG), ha sido posible aislarlo recientemente. Los anticuerpos frente
a esta proteína, marcados con sustancias fluorescentes, posibilitan
visualizar claramente los astrocitos (Bradford, 1988; Lesson, 1976).
Los astrocitos como su nombre lo sugiere, son células en forma
de estrella, con numerosas prolongaciones citoplásmicas ramificadas.
Sus núcleos son grandes, ovoides o esféricos y tienen color
pálido, con pocos gránulos finos de cromatina distribuidos
principalmente en la periferia en unión íntima con la membrana
nuclear. Los nucléolos no son patentes. El citoplasma incluye los
organelos corrientes y puede observarse claramente un centrosoma. Puede
haber pigmento lipocrómico (Lesson, 1976). Sus prolongaciones celulares
fibrosas características, así como su pericarión,
están rellenas de gliofilamentos cuyo componente proteico más
importante, llamado proteína ácida fibrilar de la glía
(PAFG), ha sido posible aislarlo recientemente. Los anticuerpos frente
a esta proteína, marcados con sustancias fluorescentes, posibilitan
visualizar claramente los astrocitos (Bradford, 1988; Lesson, 1976).
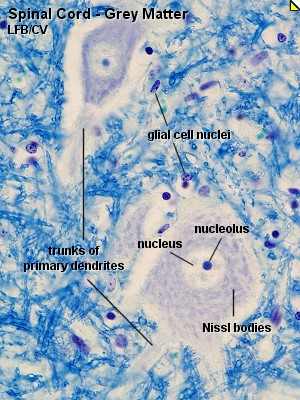
En ambos tipos las células tienen aproximadamente 8 micras; el citoplasma
perinuclear se tiñe pálidamente, por la pobreza en retículo
granuloso y en ribosomas libres, y contiene algunas mitocondrias, complejos
de Golgi, microfilamentos y, de ordinario, gran número de lisosomas
y muchas partículas de glucógeno. El núcleo es ovoide
y de forma irregular. Los astrocitos tienen gran número de prolongaciones
citoplásmicas, que se tiñen poco y contienen algunos microfilamentos.
Estas prolongaciones rodean y separan las neuronas; algunas tienen expansiones
terminales que establecen contacto con los vasos sanguíneos (pies
perivasculares o pies terminales astrocíticos) epéndimo y
piamadre en la superficie del SNC. Donde los astrocitos, o sus prolongaciones,
establecen contacto con los astrocitos vecinos o con neuronas vecinas,
puede haber desmosomas y uniones de espacio vacío. Las prolongaciones
de los astrocitos y otros elementos gliales se encuentran entre los cuerpos
neuronales, y sus axones y dendritas constituyen el neurópilo (Lesson,
1976).
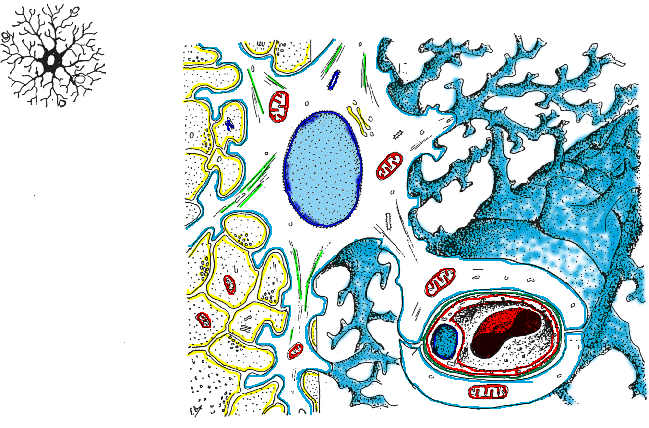 Los astrocitos pueden ser protoplasmáticos
(astrocitos tipo-1) que son los que tienen prolongaciones
ramificadas y se encuentran principalmente en la sustancia gris dispersa
entre los cuerpos neuronales; constituyen una gran proporción del
volumen citoplásmico de la sustancia gris (Bradford, 1988; Lesson,
1976). No contienen abundantes estructuras filamentosas y sus prolongaciones
tienden a ser laminares o membranosas (Lesson, 1976).
Los astrocitos pueden ser protoplasmáticos
(astrocitos tipo-1) que son los que tienen prolongaciones
ramificadas y se encuentran principalmente en la sustancia gris dispersa
entre los cuerpos neuronales; constituyen una gran proporción del
volumen citoplásmico de la sustancia gris (Bradford, 1988; Lesson,
1976). No contienen abundantes estructuras filamentosas y sus prolongaciones
tienden a ser laminares o membranosas (Lesson, 1976).
También pueden ser fibrosos (astrocitos
tipo-2), o células en forma de araña, que tienen prolongaciones
más largas y más delgadas, contienen numerosas fibrillas
citoplásmicas y están situadas principalmente en la sustancia
blanca, situados entre los haces de axones mielinizados (Lesson, 1976).
Desarrollo
de las células macrogliales
Las primeras células gliales que aparecen en el período embrionario
son las células gliales radiales (Cameron, 1991). De hecho, las
neuronas emplean estas células como orientación para encontrar
su lugar definitivo en el sistema nervioso. Al final de la gestación,
las células gliales radiales se transforman en astrocitos fibrosos.
La mayoría de los astrocitos aparecen más tardíamente
en el desarrollo, aproximadamente en el nacimiento ; su número se
va incrementando hasta después de algunas semanas de vida postnatal.
Los astrocitos que se originan durante el período embrionario se
desarrollan a partir de la zona ventricular de la placa neural, y los que
se originan postnatalmente provienen de la zona subventricular (Sánchez-Abarca,
1998).
Los oligodendrocitos surgen en período postnatal, aunque su diferenciación
o, lo que es lo mismo, la mielinización, comienza en la rata una
semana después de su nacimiento y alcanza su máximo el día
21 de vida postnatal. En el hombre, la síntesis de mielina se efectúa,
aproximadamente, desde la semana 25 de gestación hasta los 20 años
de vida adulta (Gould, 1985; Martínez, 1989).
Aunque esta pueda ser una consideración general, la realidad es
que el tiempo y el origen de los astrocitos y oligodendrocitos cambia dependiendo
del área del SNC analizada. Así, una de las cuestiones más
importantes relacionadas con el linaje glial es si comparten sus precursores
con las neuronas, es decir: derivan de una célula progenitor multipotencial
(Noble, 1991). La respuesta a esta cuestión parece depender del
área del cerebro analizada. A continuación se resume el linaje
glial en cuatro regiones diferentes del SNC:
* Desarrollo
de las células macrogliales en la corteza cerebral: en este nivel
del SNC hay un alto nivel de complejidad. La primera controversia
del linaje glial de la corteza cerebral es si las células derivan
de un precursor celular multipotencial o no (Davis y Temple, 1994; Wolswijk
y Noble, 1989). Así, algunos experimentos sugieren que la zona ventricular
de la corteza está compuesta por una mezcla de diferentes precursores
celulares. La mayoría de ellos genera un sólo tipo de células
(neuronas, astrocitos u oligodendrocitos) y el resto genera neuronas y
un sólo tipo de células de la glía. Sin embargo, cuyo
se cultivan células progenitoras de la zona ventricular, un pequeño
porcentaje de estas células son pluripotenciales, es decir, son
capaces de generar neuronas, astrocitos y oligodendrocitos (Davis y Temple,
1994).
 Estudios realizados tanto in vivo como in vitro sugieren
que en el proceso de gliogénesis las células van reduciendo
su multipotencialidad a medida que avanza el desarrollo. Así, una
vez que los precursores gliales y neuronales están diversificados,
el linaje glial puede ser seguido por el origen de tres diferentes precursores
celulares (ver figura 1). Uno de estos precursores dará lugar
a las células gliales radiales, que son las primeras en aparecer
en el SNC. Después estas células gliales radiales se transformarán
en células Bergman, células de Müller, células
ependimales. Por otro lado, los astrocitos tipo-1 se generarán a
partir de un precursor propio. Los oligodendrocitos se originarán
a partir de las células de un precursor propio(Barres et al., 1992).
Los oligodendrocitos se originarán a partir de las células
progenitoras O-2A (precursores de oligodendrocitos y astrocitos tipo-2).
Estas células progenitoras O-2A, son bipotenciales in vitro y pueden
originar, también, una segunda población de astrocitos, que
son los astrocitos tipo-2 (Wolswijk y Noble, 1989).
Estudios realizados tanto in vivo como in vitro sugieren
que en el proceso de gliogénesis las células van reduciendo
su multipotencialidad a medida que avanza el desarrollo. Así, una
vez que los precursores gliales y neuronales están diversificados,
el linaje glial puede ser seguido por el origen de tres diferentes precursores
celulares (ver figura 1). Uno de estos precursores dará lugar
a las células gliales radiales, que son las primeras en aparecer
en el SNC. Después estas células gliales radiales se transformarán
en células Bergman, células de Müller, células
ependimales. Por otro lado, los astrocitos tipo-1 se generarán a
partir de un precursor propio. Los oligodendrocitos se originarán
a partir de las células de un precursor propio(Barres et al., 1992).
Los oligodendrocitos se originarán a partir de las células
progenitoras O-2A (precursores de oligodendrocitos y astrocitos tipo-2).
Estas células progenitoras O-2A, son bipotenciales in vitro y pueden
originar, también, una segunda población de astrocitos, que
son los astrocitos tipo-2 (Wolswijk y Noble, 1989).
* Desarrollo
de las células macrogliales en el nervio óptico: el nervio
óptico contiene los axones de las células ganglionares de
la retina, las dos clases de células macrogliales (astrocitos
y oligodendrocitos) y las células microgliales (Small et al.,
1987). Los astrocitos se desarrolla a partir de células neuroepiteliales
que forman el primitivo nervio óptico, mientras que los oligodendrocitos
se desarrollan a partir de células precursoras que migran al nervio
óptico en tempranos estadios del desarrollo (Small et al.,
1987). Los oligodendrocitos derivan de precursores celulares bipotenciales
O-2A, ya que en cultivo son capaces de originar tanto oligodendrocitos
como astrocitos tipo-2, dependiendo de las condiciones de cultivo (Ffrench-Constant,
1986a; Ffrench-Constant, 1986b; Ffrench-Constant, 1986c; Raff, 1989).
 Los astrocitos aparecen en el nervio óptico de rata en el día
16 del período embrionario y van aumentando su número hasta
la sexta semana postnatal (Burne y Raff, 1997). Curiosamente, los axones
de las células ganglionares de la retina dirigen la proliferación
de los astrocitos (Raff, 1989). Dado que la muerte celular normal no parece
jugar un papel importante en el ajuste del número de astrocitos
en el nervio óptico de rata, los axones son los responsables del
número final de astrocitos, debido al control que ejercen sobre
su proliferación (Burne y Raff, 1997).
Los astrocitos aparecen en el nervio óptico de rata en el día
16 del período embrionario y van aumentando su número hasta
la sexta semana postnatal (Burne y Raff, 1997). Curiosamente, los axones
de las células ganglionares de la retina dirigen la proliferación
de los astrocitos (Raff, 1989). Dado que la muerte celular normal no parece
jugar un papel importante en el ajuste del número de astrocitos
en el nervio óptico de rata, los axones son los responsables del
número final de astrocitos, debido al control que ejercen sobre
su proliferación (Burne y Raff, 1997).
Los oligodendrocitos aparecen por primera vez después del nacimiento,
su número alcanza el máximo en el día 21 postnatal
y disminuye ligeramente en el nervio óptico adulto (Raff, 1989).
Los astrocitos tipo-2 se detectan en el nervio óptico de rata a
partir de la primera semana de vida postnatal, aunque en un número
muy pequeño.
* Desarrollo de las
células macrogliales en la retina: en la retina existe una gran
variedad de células diferentes que derivan de las células
precursoras multipotenciales . Estas células precursoras retendrán
su multipotencialidad mientras continúen dividiéndose (Turner
y Cepko, 1987). Estos precursores están capacitados para generar
tanto neuronas como fotorreceptores o células gliales Müller
(Germer et al., 1997). La única excepción son los astrocitos;
este tipo celular glial es el único no encontrado en los clones
formados a partir de los precursores multipotenciales de la retina (Watanabe
y Raff, 1988).
* Desarrollo
de las células macrogliales en la médula espinal: las células
neurales de la médula espinal se desarrollan a partir de células
progenitoras pluripotenciales. De hecho, las células neuroepiteliales
procedentes del tubo neural caudal en cultivo pueden dar origen a neuronas,
astrocitos y oligodendrocitos. Por tanto, estas células neuroepiteliales
se comportan como precursores celulares multipotenciales, capaces de generar
múltiples derivados neurales.

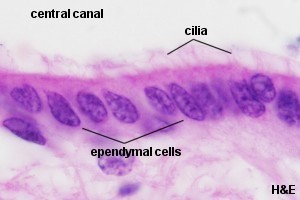 Epéndimo: Las células
ependimales revisten las cavidades del encéfalo y el conducto central
de la médula espinal. Forman una capa única de células
cúbicas o cilíndricas que poseen microvellosidades y cilias.
Las cilias son móviles y contribuyen al flujo de líquido
cefaloraquídeo.
Epéndimo: Las células
ependimales revisten las cavidades del encéfalo y el conducto central
de la médula espinal. Forman una capa única de células
cúbicas o cilíndricas que poseen microvellosidades y cilias.
Las cilias son móviles y contribuyen al flujo de líquido
cefaloraquídeo.