3.2.2.1.
Funciones de los astrocitos
Los astrocitos diferenciados tienen distintas funciones entre las que se
destacan: regulación de la composición iónica del
líquido extracelular del sistema nervioso central, inducción
de la formación de la barrera hematoencefálica (BHE), actuar
de soporte y guía de las neuronas durante la migración y
ayudar a mantener los niveles de los intermediarios del ciclo de los ácidos
tricarboxílicos en las neuronas (Kimelberg y Norenberg, 1989; Miller
y col., 1989).
 Durante el desarrollo embrionario una de las funciones principales de los
astrocitos es la de servir de soporte y guía en la migración
de las neuronas postmitóticas, así como, conducir la emisión
de prolongaciones axónicas. En el cerebro adulto, los astrocitos
realizan esta misma función durante la regeneración axonal
y la formación de nuevas sinapsis. Parece ser que entre las sustancias
implicadas en este mecanismo están las moléculas de adhesión
celular nerviosa (NCAM) y las N-caderinas (Edelman, 1983; Takeichi,
1988). En cocultivos de neuronas con astrocitos tipo 1 ó astrocitos
tipo 2 se ha demostrado que el crecimiento axonal se favorece cuando se
emplean cultivos de células neonatales en vez de células
adultas. De estas observaciones se concluye que las diferencias en la composición
molecular de las membranas astrocíticas, son las responsables del
crecimiento de las neuritas (Geisert, 1991; Smith, 1993).
Durante el desarrollo embrionario una de las funciones principales de los
astrocitos es la de servir de soporte y guía en la migración
de las neuronas postmitóticas, así como, conducir la emisión
de prolongaciones axónicas. En el cerebro adulto, los astrocitos
realizan esta misma función durante la regeneración axonal
y la formación de nuevas sinapsis. Parece ser que entre las sustancias
implicadas en este mecanismo están las moléculas de adhesión
celular nerviosa (NCAM) y las N-caderinas (Edelman, 1983; Takeichi,
1988). En cocultivos de neuronas con astrocitos tipo 1 ó astrocitos
tipo 2 se ha demostrado que el crecimiento axonal se favorece cuando se
emplean cultivos de células neonatales en vez de células
adultas. De estas observaciones se concluye que las diferencias en la composición
molecular de las membranas astrocíticas, son las responsables del
crecimiento de las neuritas (Geisert, 1991; Smith, 1993).
Los capilares cerebrales están rodeados, casi en su integridad,
por los pies terminales de las fibras astrocíticas. 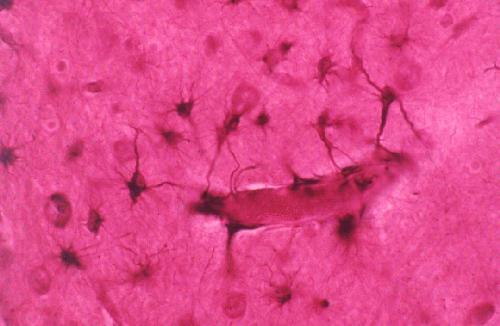 A diferencia de los que ocurre en otros tejidos, las células endoteliales
de los capilares cerebrales están fuertemente conectadas por uniones
estrechas (tight junctions). Este hecho impide el transporte paracelular
de muchas sustancias. En este sentido, los astrocitos inducen a las células
endoteliales de los capilares cerebrales a formar las uniones estrechas
y a sintetizar las enzimas carácteristicas de la barrera hematoencefálica
(Janzer y Raff, 1987).
A diferencia de los que ocurre en otros tejidos, las células endoteliales
de los capilares cerebrales están fuertemente conectadas por uniones
estrechas (tight junctions). Este hecho impide el transporte paracelular
de muchas sustancias. En este sentido, los astrocitos inducen a las células
endoteliales de los capilares cerebrales a formar las uniones estrechas
y a sintetizar las enzimas carácteristicas de la barrera hematoencefálica
(Janzer y Raff, 1987).
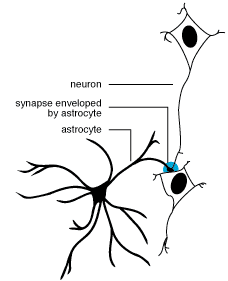
Las neuronas debido a la transmisión sinaptica liberan una serie
de neurotransmisores al medio (glutamato, aspartato, GABA, etc.). 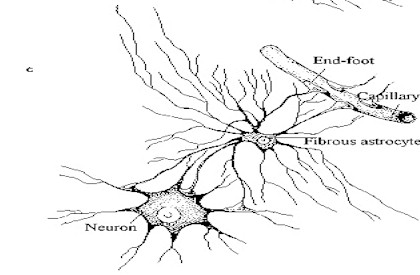 Este hecho trae consigo una disminución de la concentración
de intermediarios del ciclo de los ácidos tricarboxílicos,
principalmente oxalacetato y a-cetoglutarato,
que son fundamentalmente, los precursores de estos neurotransmisores. En
este sentido, estudios recientes han puesto de manifiesto que los astrocitos
podrían estar implicados en el mantenimiento de los niveles de los
intermediarios del ciclo de los ácidos tricarboxílicos en
las neuronas (Kaufman y Driscoll, 1992).
Este hecho trae consigo una disminución de la concentración
de intermediarios del ciclo de los ácidos tricarboxílicos,
principalmente oxalacetato y a-cetoglutarato,
que son fundamentalmente, los precursores de estos neurotransmisores. En
este sentido, estudios recientes han puesto de manifiesto que los astrocitos
podrían estar implicados en el mantenimiento de los niveles de los
intermediarios del ciclo de los ácidos tricarboxílicos en
las neuronas (Kaufman y Driscoll, 1992).
Los niveles de sodio y potasio deben estar regulados muy estrictamente
en el espacio que rodea las neuronas, de manera que pueda llevarse a cabo
los potenciales de acción. En otros tejidos, los iones externos
residen en el espacio intersticial, que en el cerebro, está ocupado
por las finas prolongaciones astrocíticas. Este hecho dió
lugar a la hipótesis llamada amortiguación
espacial de potasio (Orkand y col., 1966).  Esta hipótesis propone que los astrocitos retiran el potasio sobrante
del espacio extracelular procedente de la actividad neuronal y los transfieren
a zonas con baja concentración de potasio. La fuerza determinante
de este proceso sería el aumento local de potasio extracelular puesto
que, los astrocitos captarían el potasio debido a la alta permeabilidad
que presentan a este ión (Erecinska, 1993; Kuffler, 1966a). Más
tarde lo transmitirian de unos astrocitos a otros a través de las
llamadas uniones comunicantes (gap junctions) (Gardner-Medwin,
1986; Sáez y col., 1993). En efecto, el acoplamiento entre astrocitos
a través de las uniones comunicantes aumenta su capacidad de amortiguar
espacialmente el potasio (Mobbs y col., 1988).
Esta hipótesis propone que los astrocitos retiran el potasio sobrante
del espacio extracelular procedente de la actividad neuronal y los transfieren
a zonas con baja concentración de potasio. La fuerza determinante
de este proceso sería el aumento local de potasio extracelular puesto
que, los astrocitos captarían el potasio debido a la alta permeabilidad
que presentan a este ión (Erecinska, 1993; Kuffler, 1966a). Más
tarde lo transmitirian de unos astrocitos a otros a través de las
llamadas uniones comunicantes (gap junctions) (Gardner-Medwin,
1986; Sáez y col., 1993). En efecto, el acoplamiento entre astrocitos
a través de las uniones comunicantes aumenta su capacidad de amortiguar
espacialmente el potasio (Mobbs y col., 1988).
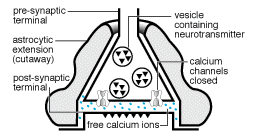
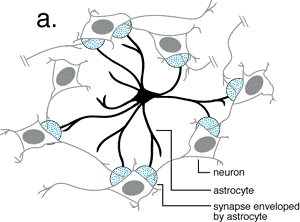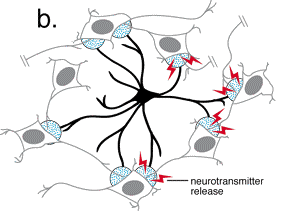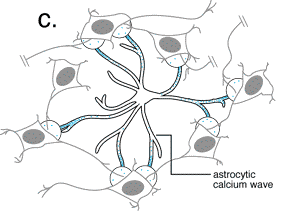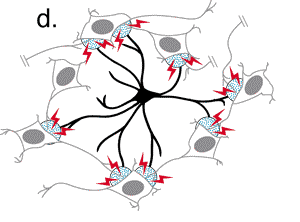 Recientemente se ha comenzado a conjeturar sobre la posibilidad de que
los astrocitos tengan una función mucho más activa en el
SNC y no un papel meramente protector de las neuronas. Se ha propuesto
que los astrocitos presentan un tipo de excitabilidad basada directamente
en la dinámica del ión calcio intracelular y que es esencialmente
independiente del potencial de membrana (Cornell-Bell y Finkbeiner, 1991;
Cornell-Bell y col., 1990). Así, en los astrocitos se expresan una
amplia variedad de receptores funcionales para agentes neuroactivos (Dermietzel,
1991a; Cornell-Bell y col., 1990; Jensen, 1990; Salm y McCarthy, 1990).
Recientemente se ha comenzado a conjeturar sobre la posibilidad de que
los astrocitos tengan una función mucho más activa en el
SNC y no un papel meramente protector de las neuronas. Se ha propuesto
que los astrocitos presentan un tipo de excitabilidad basada directamente
en la dinámica del ión calcio intracelular y que es esencialmente
independiente del potencial de membrana (Cornell-Bell y Finkbeiner, 1991;
Cornell-Bell y col., 1990). Así, en los astrocitos se expresan una
amplia variedad de receptores funcionales para agentes neuroactivos (Dermietzel,
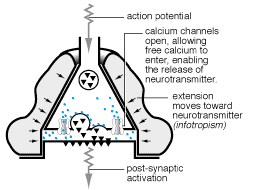
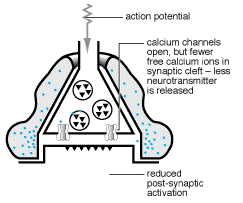
1991a; Cornell-Bell y col., 1990; Jensen, 1990; Salm y McCarthy, 1990). 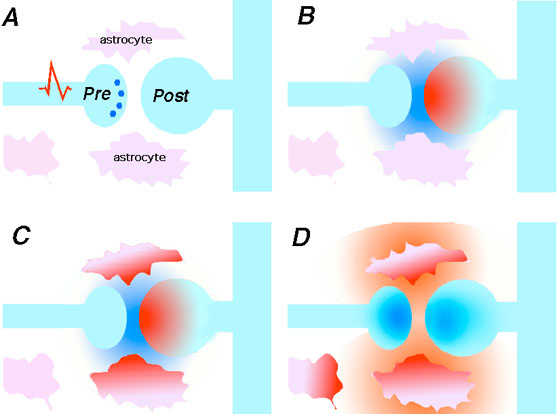 En estos estudios se ha establecido que el glutamato y otros neurotransmisores
pueden provocar oscilaciones en los niveles del ión calcio intracelular
y la propagación de estas ondas de calcio, creando en definitiva
una forma de exitabilidad, basada en las corrientes de Ca+2
(Cornell-Bell y col., 1990; Jensen y Chiu, 1991b;
Jensen y col., 1991a). Esta forma de excitabilidad comienza con la liberación
de iones calcio de los depositos intracelulares (Cornell-Bell y Finkbeiner,
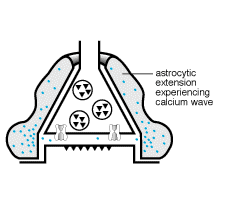
1991) y continua con la oscilación en los niveles de calcio, que
se propaga dentro del astrocito exitado a los astrocitos adyacentes, a
través de las uniones comunicantes. Recientemente, se ha demostrado
que las inervaciones aferentes de las neuronas glutaminérgicas provocan
la formación de las ondas de calcio en los astrocitos (Dani y col.,
1992; Nedergaard, 1994).
En estos estudios se ha establecido que el glutamato y otros neurotransmisores
pueden provocar oscilaciones en los niveles del ión calcio intracelular
y la propagación de estas ondas de calcio, creando en definitiva
una forma de exitabilidad, basada en las corrientes de Ca+2
(Cornell-Bell y col., 1990; Jensen y Chiu, 1991b;
Jensen y col., 1991a). Esta forma de excitabilidad comienza con la liberación
de iones calcio de los depositos intracelulares (Cornell-Bell y Finkbeiner,
1991) y continua con la oscilación en los niveles de calcio, que
se propaga dentro del astrocito exitado a los astrocitos adyacentes, a
través de las uniones comunicantes. Recientemente, se ha demostrado
que las inervaciones aferentes de las neuronas glutaminérgicas provocan
la formación de las ondas de calcio en los astrocitos (Dani y col.,
1992; Nedergaard, 1994).
Oligodendroglía