3.1.1.2.
Clasificación funcional.
En el ámbito
funcional las neuronas se clasifican en sensoriales, motoras o interneuronas.
Las neuronas sensoriales conducen impulsos desde los receptores hasta el
cerebro y la médula espinal; estos impulsos son informativos (visión,
sonido, tacto, dolor, etc.). Estas neuronas son
los componentes sensoriales aferentes de los nervios espinales y craneales;
sus cuerpos celulares forman en gran parte la médula espinal (raíz
posterior) y los ganglios craneales. Generalmente este tipo de neuronas
posee una estructura de tipo seudounipolar o bipolar (Meyer, 1985).
Las motoneuronas
conducen el impulso desde el cerebro y la médula espinal hasta los
efectores (músculos y glándulas) lo que origina la contracción
de las fibras musculares o la secreción glandular. Estas neuronas
son el componente motor eferente de los nervios espinales y craneales.
Por lo general su estructura es de tipo multipolar (Roselli, 1997).
Existe otro tipo de neuronas cuyos somas y procesos permanecen en el SNC
y se les conoce como interneuronas las cuales no tienen contacto
directo con estructuras periféricas (receptores y efectores).
Existe un grupo importante de interneuronas cuyos axones descienden y terminan
en motoneuronas en el tronco del encéfalo y en la médula
espinal; a estas células se les denomina motoneuronas altas. Las
interneuronas son responsables de la modificación, coordinación,
integración, facilitación e inhibición que debe ocurrir
entre la entrada sensorial y la salida motora. Por lo general su estructura
es de tipo multipolar (Meyer, 1985).
3.1.1.3.
Clasificación por tipo de neurotransmisor.
Neuronas
colinérgicas
Estas neuronas
utilizan la acetilcolina (Ach) como neurotransmisor químico, la
cual se encuentra en los terminales de las placas motoras, que son las
uniones entre el nervio y el músculo estriado, haciendo posible
la contracción de diferentes músculos y la estimulación
de las glándulas exocrinas. Para ejercer su acción posee
dos tipos de receptores específicos: muscarínicos, localizados
en el músculo liso y cardíaco, son bloqueados por la atropina
y nicotínicos, localizados en ganglios autónomos y en la
unión neuromuscular, son bloqueados por el hexametonio y el curare
(Rosselli, 1997).
Neuronas
catecolaminérgicas
Dentro de este
tipo de neuronas se identifican principalmente dos, de acuerdo al neurotransmisor
que posean: noradrenalina (NA) o dopamina (DA). Las neuronas que
utilizan como mensajero químico la noradrenalina, actúan
bajo la excitación generalizada del sistema simpático por
la emoción y el ejercicio, produciendo efectos cardiovasculares:
vasoconstricción y estimulación cardíaca. La transmisión
noradrenérgica tiene lugar en las sinapsis posganglionares simpáticas
en el músculo liso, músculo cardíaco y glándulas
exocrinas. La noradrenalina actúa sobre dos tipos de receptores
adrenérgicos: alfa y beta (Meyer, 1985). Las neuronas que utilizan
dopamina como neurotransmisor provocan reacciones cardiovasculares similares
a la de la adrenalina, estas acciones tienen lugar por la activación
de los receptores beta (vasodilatación y estimulación cardíaca)
y alfa (vasoconstricción), además, presentan propiedades
específicas como el incremento del débito renal, el flujo
mesentérico y el coronario, acciones que se deben a la activación
de receptores dopaminérgicos denominados D, que están asociados
a la adenilciclasa (Bradford, 1988).
Neuronas
indolaminérgicas
Este tipo de
neuronas utiliza la serotonina como agente químico que actúa
en la sinapsis neuronal. Este neurotransmisor se encuentra ubicado especialmente
en las neuronas del tronco cerebral en la región del rafé
medio del puente y del mesencéfalo. Interviene en varios tipos de
regulación: mantenimiento del estado anímico, regulación
de la temperatura, analgesia, conducta sexual, agresividad, control de
los reflejos monosinápticos y del tono muscular e inhibición
del tono simpático. Los receptores específicos de la serotonina
son bloqueados por la dietilamina del ácido lisérgico y la
metisergida (Meyer, 1985).
Neuronas
adrenérgicas
Utilizan como
neurotransmisor la adrenalina, se encuentran en la porción rostral
del bulbo raquídeo, sus axones ascienden gasta el hipotálamo
o descienden a la médula espinal; al igual que las neuronas noradrenérgicas
actúan sobre receptores adrenérgicos alfa y beta (Bradford,
1988).
Neuronas
GABAérgicas
Dichas neuronas transmiten el impulso nervioso mediante el GABA, el cual
es una aminoácido inhibitorio. Están ubicadas en el cerebelo,
cuernos dorsales de la médula, retina, hipocampo y el hipotálamo,
funcionan como transmisor inhibidor en el sistema nervioso de vertebrados
e invertebrados (Barr, 1994).
Otras
Existen diversos neurotransmisores utilizados por las neuronas para conducir
el impulso nervioso, dando diferentes nombres a las mismas. Dentro de estos
encontramos: el glutamate (Glu) y el aspartato (Asp) considerados
excitadores del sistema nervioso, la glicina (Gly) y la taurina considerados
como inhibidores.
El
Grupo de Neuronas
Recientemente existe un gran interés en enfocar el estudio de la
mente intentando construir una teoría global que partiendo desde
la biología conecta procesos cerebrales con procesos mentales. Uno
de los autores que ha afrontado esta tarea es G. M. Edelman, premio
Nobel de medicina en 1972. Edelman considera que la unidad básica
de procesamiento del cerebro es el grupo de neuronas, y presenta una teoría
que nos permite, al menos, iniciarnos en como surgen los pensamientos desde
el cerebro. Veámosla brevemente.
Edelman concibe el cerebro como un sistema selectivo, en el que
la selección opera durante el tiempo de vida del sistema.
Para sobrevivir, un organismo debe o heredar o crear criterios que le permitan
clasificar el mundo en categorías perceptuales de acuerdo con sus
necesidades adaptativas. Además el mundo, incluso para el tiempo
de vida de un organismo, está lleno de novedad, lo que exige que
estos procesos de categorización puedan reestructurarse, renovarse
y reiniciarse continuamente. El mundo, para el organismo, no se da por
completo de una vez, sino que se construye en un proceso constante y continuo.
Por tanto, los órganos que se encargan de estas tareas, es decir,
el cerebro en último término, debe ser flexible, pero también,
como consecuencia, único. Esto está de acuerdo con la enorme
variación funcional y estructural en muchos niveles: molecular,
celular, anatómico, fisiológico y conductual, que muestran
los sistemas nerviosos por lo que, a pesar de las semejanzas en los individuos
de una especie, el grado de variación individual de cerebro en cerebro
excede lo que podía tolerarse en un proceso de fabricación
de ingeniería. Visto así, cualquier teoría interesante
sobre la mente tendrá que tener en cuenta estas observaciones y
no podrá generalizar, a menos que contemple en la descripción
estructural, orgánica, las fuertes diferencias y la exigencia de
flexibilidad y variación que impone la novedad del mundo. Por eso
Edelman rechaza las teorías que contemplan el cerebro como un sistema
que procesa información.
A partir de aquí Edelman enuncia su tesis fundamental, a
saber, que el cerebro es un sistema selectivo, en el que la selección
opera durante el tiempo de vida del individuo. Para el desarrollo de esta
tesis fundamental elabora lo que él denomina la
teoría de la selección del grupo de neuronas (TNGS).
La TNGS es una teoría de poblaciones que postula precisamente que
la habilidad de los organismos para categorizar un mundo no etiquetado
y para comportarse en él de una manera adaptativa surge no de la
transferencia de instrucciones o de información sino de procesos
de selección bajo variación. La TNGS considera que hay una
generación continua de diversidad en el cerebro. En el cerebro embrionario,
hay variación y selección en la migración de poblaciones
celulares y durante la muerte de células. También en la formación
de las sinapsis. Y en el cerebro maduro, en la amplificación diferencial
de la eficacia de las sinapsis.
Esto tiene como consecuencia la formación de grupos neuronales y
que el proceso es modificado continuamente por reentradas de señales.
Veamos esto más despacio.
Postulados
Básicos de la TNGS
La TNGS propone tres mecanismos para responder de la producción
de conductas adaptativas por parte de los organismos con sistemas nerviosos
complejos: selección en el desarrollo, selección en la experiencia
y reentradas de señales.
Cada uno de estos mecanismos actúa dentro y entre colectivos que
consisten en cientos de miles de neuronas fuertemente interconectadas denominadas
grupos de neuronas.
Además la teoría propone que la selección a través
de la amplificación sináptica diferencial está restringida
por la acción de sistemas de valor derivados evolutivamente: sistemas
neuromodulatorios dotados con proyecciones difusas que señalan el
posible valor adaptativo para el organismo en su totalidad de la ocurrencia
de ciertos eventos.
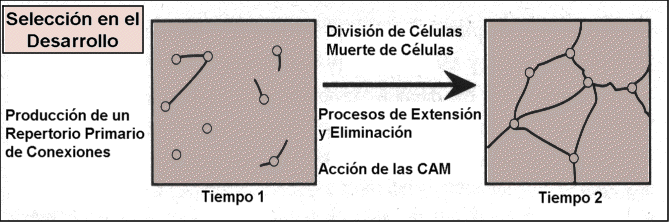
1.
Variación y Selección en el desarrollo.
La diversidad estructural del sistema nervioso y los detalles de la neuroanatomía
no están estrictamente programados por el código genético.
Esta diversidad surge durante el desarrollo en la regulación epigenética
dinámica de la división, adhesión, migración
y muerte de la célula, así como de la actividad neural misma.
Durante la producción del sistema nervioso se van creando neuronas
y agrupaciones de células que permanecerán o no dependiendo
del refuerzo que otorgue la experiencia. La adhesión y migración
son gobernadas por unas series de moléculas morforeguladoras llamadas
CAMs (moléculas de adhesión célular) y SAMs
(moléculas de adhesión de sustratos).
Esto lleva a la formación de repertorios primarios dentro de regiones
anatómicas dadas que contienen un gran número de grupos de
neuronas o circuitos locales.
 2.
Selección en la experiencia.
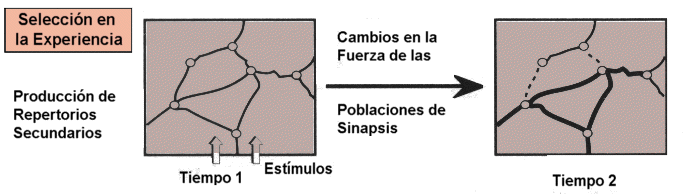
2.
Selección en la experiencia.
Después de que la mayoría de las conexiones anatómicas
de los repertorios primarios se han establecido, las actividades de los
grupos de neuronas que funcionan particularmente continúan siendo
dinámicamente seleccionadas por mecanismos de cambios sinápticos
subsiguientes dirigidos por la conducta y la experiencia.
Será la experiencia del organismo la que tenderá a reforzar
algunos de los circuitos que se han establecido en la fase anterior dentro
del grupo y entre grupos, otros tenderán a desaparecer si el organismo
no los requiere con la frecuencia que indicará su utilidad. De esta
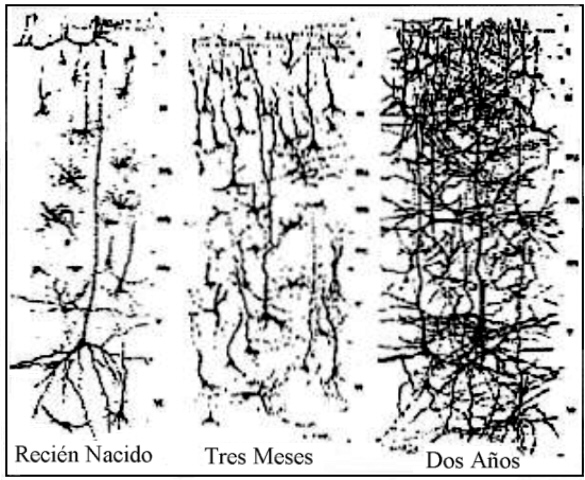
manera la maraña de conexiones que encontramos en un individuo de
dos años se irá simplificando para consolidar las conexiones
útiles dependiendo del tipo de experiencia que realice el organismo.
La selección en la experiencia lleva finalmente a la formación
de repertorios secundarios de grupos neurales como respuesta a patrones
particulares de señales.
 3.
Reentradas de señales.
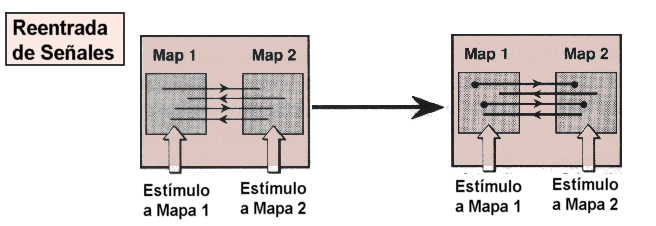
3.
Reentradas de señales.
La selección en la experiencia conlleva correlaciones de señales
estadísticas entre grupos de neuronas pre y postsinápticas,
mejor que la transmisión de mensajes codificados de una neurona
a otra.
Si estas señales han de ser adaptativas tendrán que reflejar
las señales que surjan en el mundo real. Esto se realiza señalando
reentradas en y entre mapas neuronales. Estos recorridos neurales que relacionan
hojas de receptores sensoriales con registros particulares del sistema
nervioso central proveen un medio de reforzar regularidades espacio-temporales.
Una reentrada puede definirse como una señalización paralela
continua entre grupos de neuronas separadas que ocurre a lo largo de conexiones
anatómicas ordenadas de manera bidireccional y recursiva. Es, pues,
un proceso dinámico que es inherentemente paralelo y distribuido
y que debe diferenciarse de la retroalimentación. Las reentradas
no tienen una dirección preferida y no tiene una función
de input o output predefinido.
 Una
de las premisas fundamentales de la TNGS es, entonces, que la coordinación
selectiva de patrones complejos de interconexiones entre grupos de neuronas
por medio de reentradas es la base de la conducta. Para la teoría
de Edelman, la reentrada es la base principal para poder trazar el puente
entre la fisiología y la psicología.
Una
de las premisas fundamentales de la TNGS es, entonces, que la coordinación
selectiva de patrones complejos de interconexiones entre grupos de neuronas
por medio de reentradas es la base de la conducta. Para la teoría
de Edelman, la reentrada es la base principal para poder trazar el puente
entre la fisiología y la psicología.
Este puente comienza a realizarse cuando múltiples mapas que están
conectados entre sí por doble entrada a la conducta sensomotor del
organismo comienzan a emparejar sus outputs creando un mapa global que
da lugar a respuestas categoriales perceptivas.
4.
Categorización Perceptiva:
La discriminación selectiva de un objeto o evento de otros objetos
o eventos con propósitos adaptativos se produce con lo que Edelman
denomina Pareja de Clasificación.
Pareja de Clasificación:
Es una unidad mínima que consiste en dos mapas funcionales diferentes
conectados por doble entrada. Si, durante cierto periodo de tiempo, reentradas
específicas conectan ciertas combinaciones de grupos de neuronas
de un mapa con otras combinaciones en el otro, las funciones y actividades
en un mapa se conectan y correlacionan con las del otro mapa.
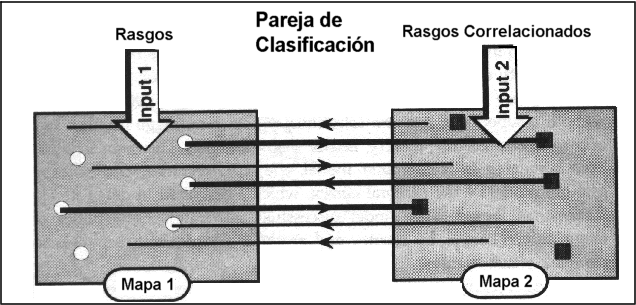
Si los mapas en cuestión están conectados topográficamente,
entonces correlacionan acontecimientos en una localización espacial
en el mundo.
Este mapa global asegura la creación de un bucle dinámico
que continuamente coteja los gestos y posturas del organismo con el muestreo
independiente obtenido de varias clases de señales sensoriales.
El mapa global permite interaccionar con partes no concatenadas del cerebro
(hipocampo, el ganglio basal y el cerebelo) en la medida en que estas estructuras
están conectadas con mapas locales mediante múltiples reentradas.
Por supuesto, para la categorización perceptiva, que iniciará
los procesos superiores y de conciencia, hay que suponer como condición
algún sistema de valor que se ha ido produciendo a lo largo de la
evolución de la especie.
Edelman denomina valor a las estructuras
fenotípicas que reflejan la selección evolutiva principal
y que contribuyen a la conducta adaptativa y a la construcción del
fenotipo. Estos sistemas de valor podrían percibir la ocurrencia
de conductas adaptativas y seleccionarían los eventos neuronales
que las producen en función de los valores simples que contengan.
Estos valores pueden expresarse en proposiciones tales como: "Comer es
mejor que no comer" o "ver es mejor que no ver".
Los procesos que producen estos mapas globales (con sus patrones asociados
de selección del grupo de neuronas y de cambio sináptico)
crean una representación espaciotemporal continua de objetos o de
eventos.
Dentro de estos procesos globales, los cambios de gran alcance en la fuerza
sináptica tienden a favorecer la actividad mutua de reentradas de
aquellos grupos cuya actividad ha sido correlacionada a través de
diferentes mapas durante la conducta pasada. Tales cambios sinápticos
proveen la base para la memoria.
De esta manera los recuerdos en los mapas globales no son almacenados,
fijados o codificados de tal manera que puedan invocarse y recuperarse
siempre de la misma manera como haríamos con los registros de un
disco duro de nuestros ordenadores. En lugar de ello, la memoria resulta
de un proceso de recategorización continua, que, por su naturaleza,
debe ser procedural y que debe conllevar una actividad motora continua
y una repetición frecuente.
Con esto, categorización y memoria, obtenemos la condición
necesaria para el aprendizaje. Edelman consigue un paso más
en su teoría cuando incluye, además de los procesos de categorización
y memoria, enlaces sinápticos entre los procesos que producen los
mapas globales y la actividad de los centros hedonistas y el sistema límbico
de manera que satisfacen nuestras necesidades homeostáticas, apetitivas
y consumatorias.
El aprendizaje se entiende entonces como la satisfacción de necesidades
del organismo, necesidades que han creado expectativas al asociar estas
necesidades con nuestras categorizaciones.
Hasta aquí una explicación estrictamente neurocientífica
de los procesos iniciales por los cuales iniciamos nuestra representación
del mundo. Quizá resulte un poco complejo, pero en la continuación
del tema tendremos tiempo para ir integrando toda la explicación.
Resumen
Gráfico
1. El cerebro en su
fase embrionaria produce muchas neuronas, más de las que necesitará
posteriormente.
2. La neuronas que
se vean reforzadas por la experiencia y conducta del individuo empezarán
a establecer conexiones entre sí.
3. Un mecanismo de
refuerzo opera igualmente entre las conexiones establecidas. Así
unas se consolidarán y otras se debilitarán hasta desaparecer.
Este refuerzo viene igualmente determinado por la experiencia del organismo.
4. Un mecanismo de
reentrada se establecerá entre los grupos de neuronas conexionados
entre sí que permitirán la construcción de mapas locales
y después globales, que constituyen la base para la formación
de imágenes mentales en el cerebro.

Clases
de Neuroglía
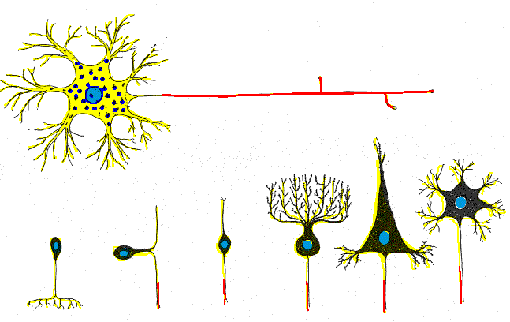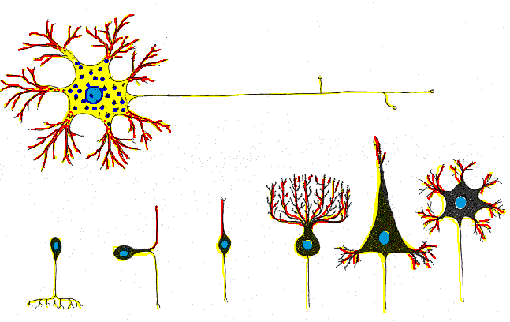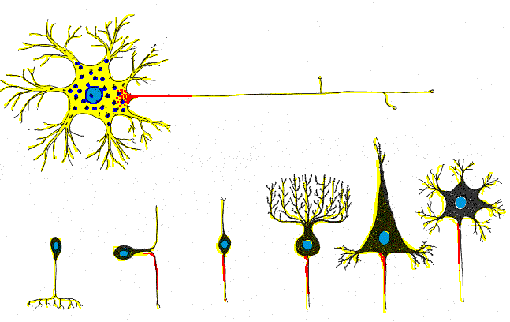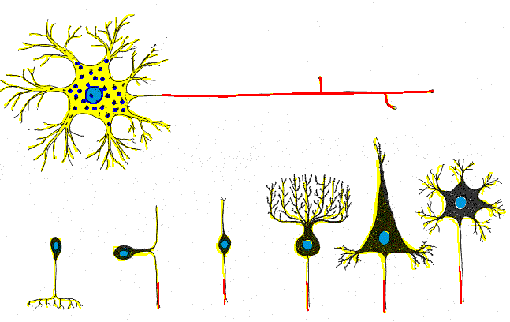 Algunas neuronas solo tienen dos procesos fundidos que en ocasiones parecen
uno, se les llama seudounipolares, las cuales están restringidas
a grupos de cuerpos celulares neuronales localizados en los ganglios, en
este tipo de neuronas los procesos cortos se ramifican del soma y
se dividen en un proceso central que conducen impulsos hacia el cuerpo
(Barr, 1994).
Algunas neuronas solo tienen dos procesos fundidos que en ocasiones parecen
uno, se les llama seudounipolares, las cuales están restringidas
a grupos de cuerpos celulares neuronales localizados en los ganglios, en
este tipo de neuronas los procesos cortos se ramifican del soma y
se dividen en un proceso central que conducen impulsos hacia el cuerpo
(Barr, 1994).
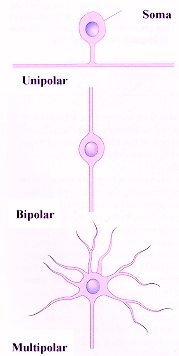
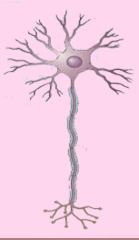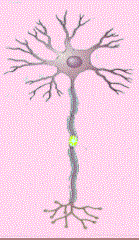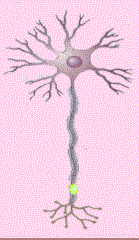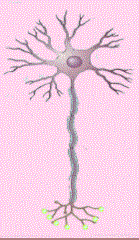
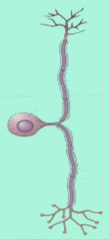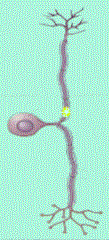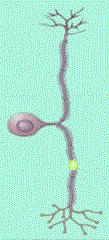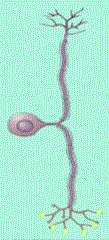
 Las neuronas multipolares se caracterizan por poseer un axón y dos
o más dendritas. Son comunes en el sistema nervioso central.
Las neuronas denominadas Golgi son células multipolares cuyos axones
se extienden hacia distancias considerables hasta llegar a la célula
Diana. Algunos ejemplos son las células piramidales de la corteza
cerebral, las células de Purkinje del cerebelo y las células
del asta anterior de la médula espinal. Las neuronas multipolares
que tienen axón corto que termina cerca del cuerpo celular de donde
se origina se llaman neuronas de Golgi II; existen como células
estrelladas o granulares de la corteza cerebral (Cooper, 1994).
Las neuronas multipolares se caracterizan por poseer un axón y dos
o más dendritas. Son comunes en el sistema nervioso central.
Las neuronas denominadas Golgi son células multipolares cuyos axones
se extienden hacia distancias considerables hasta llegar a la célula
Diana. Algunos ejemplos son las células piramidales de la corteza
cerebral, las células de Purkinje del cerebelo y las células
del asta anterior de la médula espinal. Las neuronas multipolares
que tienen axón corto que termina cerca del cuerpo celular de donde
se origina se llaman neuronas de Golgi II; existen como células
estrelladas o granulares de la corteza cerebral (Cooper, 1994).
