|
 |
 |
|
|||
|
RESEARCHThe Quest for Perfect TimingMolecular intricacies of animal model clocks provide insight to humans
Researchers have pondered, and investigated, for decades
why one person is alert and productive at 6 a.m. while another can't even
focus before noon.1 But now, their persistence is paying off:
chronobiologists, those who investigate circadian rhythms, or daily
clocks, are finally making concrete links between sleep patterns in humans
and a menagerie of well-studied animal models.
As with many behavioral studies, it's the extreme or unusual cases that eventually inform scientists about normal processes. A few years ago, a woman complained to investigators at the University of Utah in Salt Lake City that she felt an overwhelming desire to fall asleep around 7:30 p.m., and wake up before dawn, around 4:30 a.m. And she was not the only one--other members in her family had the same problem. This started Utah investigators on a research project that culminated in the first study to link a human genetic syndrome to what others had been discovering in animal clock-gene investigations. These family members, eventually recruited into the study, suffer from familial advanced sleep phase syndrome (FASPS). Earlier this year, senior authors Louis Ptacek, a Howard Hughes Medical Institute (HHMI) investigator and a professor in the departments of neurology and human genetics at Utah, and university colleague Ying-Hui, a research associate professor of neurobiology and anatomy, published their study describing the exact mutation involved in FASPS.2 People affected by FASPS have a mutation within a kinase-binding region of the hPer2 gene near the telomere of chromosome 2. And, it turns out that hPer2 is a human homologue of the period gene found in fruit flies, which encodes a key protein in the Drosophila time-keeping mechanism. But understanding why this connection is so significant requires a look at the genetics of the fruit fly's internal clock. Fruit Fly Chronobiology 101
What makes the Utah study especially interesting, says
Michael Young, a genetics professor at Rockefeller
University in New York and director of its National Science Foundation
Center for Biological Timing, is what it uncovers about the regulation of
normal human wake-sleep cycles. "In the beginning there was
period," he quips. The fruit fly gene period was cloned in
his lab and that of Michael Rosbash and Jeffrey Hall at Brandeis University in 1984. In the
1990s, a series of genes and proteins that make up the fruit fly clock
were also cloned. So far, seven of these have been characterized in a
number of labs3: period, timeless, clock,
cycle (Bmal-1 in mammals), double-time (which encodes
casein kinase 1Î, shaggy (GSK-3 in
mammals), and vrille.415
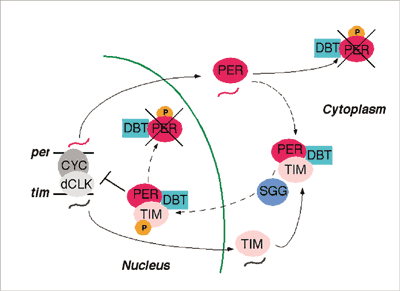
Central to the molecular mechanism of the fly's clock is the diurnal production, movement into the nucleus, and breakdown of the two-protein complex of TIMELESS and PERIOD. The transcription factors CLOCK, CYCLE, and VRILLE control production; these factors flip the genetic switch on and off for ultimately making TIMELESS and PERIOD proteins. Added to the mix are the kinases DOUBLE-TIME and SHAGGY, which are enzymes that attach phosphate groups to PERIOD and TIMELESS, respectively. When the period and timeless genes are first activated, TIMELESS proteins start to accumulate, but PERIOD proteins hold off a bit. Initially, PERIOD proteins bind with the kinase DOUBLE-TIME because it's always abundant in cells. This partnership leads to PERIOD'S phosphorylation and degradation, which continues all day long. By early evening, TIMELESS proteins become so plentiful that PERIOD has two potential partners--TIMELESS and DOUBLE-TIME. When PERIOD binds with TIMELESS, it's protected from DOUBLE-TIME and therefore from breakdown. DOUBLE-TIME still binds to PERIOD, and eventually the now three-protein complex makes it into the nucleus. Here the complex signals the cell to start decreasing production of PERIOD and TIMELESS proteins. This process finally ends at sunrise. Light-sensitive TIMELESS is eliminated from the cell, leaving PERIOD and DOUBLE-TIME alone once again. This in turn leads to the phosphorylation and breakdown of PERIOD, and the cycle starts anew. Essentially, says Young, "it's a very delicate balance between the two kinases to set the right rate of the clock." The phosphorylation of TIMELESS by SHAGGY has the opposite effect of what DOUBLE-TIME does to PERIOD protein. PERIOD's phosphorylation makes the clock go slower early in the cycle by removing PERIOD protein and SHAGGY phosphorylation of TIMELESS, then makes the clock go faster by pushing the protein complex into the nucleus more quickly. A mutation in the casein kinase 1Î binding site of the hPER2 protein somehow advanced the clocks of the FASPS family. Utah's Fu theorized how this might work, considering the scenario worked out in flies. "Our working hypothesis is that usually hPER2 is highly regulated by CK1Î through phosphorylation," he says. "In the normal case, CK1Î will just phosphorylate PERIOD and make it degrade really fast." With the mutation, the hPER2 protein becomes a poor substrate for CK1Î, causing fewer phosphates to be added. This makes the hPER2 protein more stable than in the normal cell. The hPER2 protein accumulates faster, enters the nucleus prematurely, thereby advancing the internal clock. Although the clock genes present in flies have human homologues, salient differences exist. In mammals, the clock is more complex, having more of each type of genethree periods, for example. Then, rather than TIMELESS being PERIOD's partner in the protein complex, it seems most likely that a mammalian protein called cryptochrome is PERIOD's partner. In the fly, cryptochrome is a photoreceptor for the clock, connecting the clock to light from the surrounding environment, but in a different way. In fact, Amita Sehgal, associate professor of neuroscience and an HHMI assistant investigator at the University of Pennsylvania School of Medicine (and a former postdoc with Young) is currently looking into cryptochrome's role in the breakdown of the TIMELESS protein. Jump to MammalsIt wasn't until the late 1990s that researchers cloned
mammalian clock genes. "There are now at least eight that we know are
intimately involved in the internal clockwork," says Steve Reppert, chairman of neurobiology at the
University of Massachusetts in Worcester. Reppert's lab at the
Massachusetts General Hospital, where he is also a professor at Harvard
Medical School, was involved in figuring out that mammalian
timeless doesn't perform the same role as timeless in fruit
flies and that the cryptochromes have taken over timeless functions
in mammals.16,17
The master clock in mammals is located in the suprachiasmatic nucleus (SCN), a group of about 10,000 cells in the hypothalamus at the brain's base. About five years ago, Reppert's lab began to discover how the SCN clock was put together at the molecular level. They found that the several feedback loops in mammals are very similar as to what is proposed in flies.18 The SCN receives direct enervation from the retina, and the light-dark cycle is the primary way that the clock is coordinated to a 24-hour day. In May, Reppert's group described how the three period genes in mammals function in the SCN clock.19 "The bottom line is that the Per1 and Per2 genes have nonoverlapping, essential functions for the clockwork, while Per3 is not necessary," he says. These incremental discoveries about how the SCN works at a molecular level might one day help in understanding the sleep problems of Alzheimer's disease patients. In another recent study, David Harper, assistant director of research at the geriatric psychiatry program at McLean Hospital in Belmont, Mass., which is part of the Harvard Medical School, tracked core body temperature and activity levels of 38 elderly people with probable Alzheimer's disease (AD). After they died, he examined their brain tissue and found that 11 of them had varying types of non-AD dementia diseases of the fronto-temporal region of the brain.20 "What was surprising was when we saw the data for the non-AD patients, they had a normal pattern of core body temperature, but the rest activity was entirely disorganized," he says. "It almost looked like they had SCN lesions. The activity for these patients looked like their circadian system wasn't working, but their core body temperature regulation was. But in AD patients, it looked like the circadian system was working but was unable to keep the correct time." Core body temperature is a direct measurement of activity in SCN cells; whereas, rest activity measures movement of patients every five minutes and is dependent on many factors. Knowing exactly how studies like Utah's genetic investigation of FASPS and basic research in model animals will affect sleep disorder treatments is still far off, says Ptacek. "The simple answer is that we can't really manipulate the system. We can't adjust the clock unless we understand it better. But our hope is that in understanding the ticking of the clock, the working of those gears, then we'll be able to test compounds in combinatorial libraries, and maybe design rational strategies from this knowledge." References
1. R. Lewis, "Chronobiology researchers say their field's time has come," The Scientist, 9[24]:14, Dec. 11, 1995. 2 K.L. Toh et al., "An hPer2 phosphorylation site mutation in familial advance sleep phase syndrome," Science, 291:10403, 2001. 3. Clock and cycle were isolated independently by Rosbash and Hall; Steven Kay from the Scripps Research Institute; and Isaac Edery from Rutgers University. The remaining genes were described by Young's lab; and in the case of timeless in collaboration with Amita Sehgal from the University of Pennsylvania and Charles Weitz from Harvard Medical School. 4. J.E. Rutila et al., "CYCLE is a second bHLH-PAS clock protein essential for circadian rhythmicity and transcription of Drosophila period and timeless," Cell, 93:80514, 1998. 5. R. Allada et al., "A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless," Cell, 93:791804, 1998. 6. T. K. Darlington et al., "Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim," Science, 280:1599603, 1998. 7. K. Bae et al., "Circadian regulation of a Drosophila homolog of the mammalian Clock gene: PER and TIM function as positive regulators," Molecular Cell Biology, 18:614251, 1998. 8.M. Myers et al., "Positional cloning and sequence analysis of the Drosophila clock gene timeless," Science 270, 8058, 1995. 9. A. Sehgal et al. "Rhythmic expression of timeless: A basis for promoting circadian cycles in period gene autoregulation," Science, 207:80811, 1995. 10. N. Gekakis et al., "Isolation of timeless by per protein interaction: Defective interaction between timeless protein and long-period mutant perl," Science, 207:81115, 1995. 11. J.L. Price et al., "double-time is a novel Drosophila clock gene that regulates period protein accumulation," Cell, 94:8395, 1998. 12. B. Kloss et al., "The Drosophila clock gene double-time encodes a protein closely related to human casein kinase 1Î, Cell, 94:97107, 1998. 13. N. Nadoo et al., "A role for the proteosome in the light response of the timeless clock protein, Science, 285:173741, 1999. 14. S. Martinek et al., "A role for the segment polarity gene shaggy/GSK-3 in the Drosophila circadian clock," Cell, 105:76979, 2001. 15. J. Blau and M.W. Young, "Cycling vrille expression is required for a functional Drosophila clock," Cell, 99:66171, 1999. 16. A.L. Gotter et al., "A time-less function for mouse Timeless," Nature Neuroscience, 3: 7556, 2000. 17. K. Kume et al., "mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop," Cell, 98: 193205, 1999. 18. L.P. Shearman et al., "Interacting molecular loops in the mammalian circadian clock," Science, 288:10139, 2000. 19. K. Bae et al., "Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock," Neuron, 30, 52536, 2001. 20. D.G. Harper et al., "Differential circadian rhythm disturbances in men with Alzheimer disease and frontotemporal degeneration," Archives of General Psychiatry, 58(4):35360, 2001.
© Copyright 2001, The Scientist, Inc. All rights
reserved. We welcome your opinion. If you would like to comment on this article, please write us at mailto:editorial@the-scientist.com?Subject=The Scientist, Jul. 9, 2001, The Quest for Perfect Timing News | Opinions & Letters | Research | Hot Papers LabConsumer | Profession | About The Scientist | Jobs Classified | Web Registration | Print Subscriptions | Advertiser Information |