4.2.3. Señalización sináptica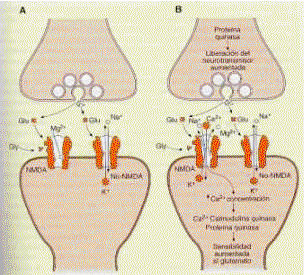
La sinapsis es el proceso esencial en la comunicación neuronal y constituye el lenguaje básico del sistema nervioso. Afortunadamente, las semejanzas de los mecanismos sinápticos son mucho más amplias que las diferencias, asociadas éstas a la existencia de distintos neurotransmisores con características particulares.
Elliot en 1904 fue el primero que sugirió la posibilidad de que la información era transferida de una neurona a otra por la liberación de una sustancia química desde las fibras nerviosas; Loewi es, sin embargo, el primero que mostró la existencia de una sustancia química en el líquido perfundido con la estimulación del nervio vago y fue su colaborador Navratil quien más tarde demostró que esta sustancia era la acetilcolina.
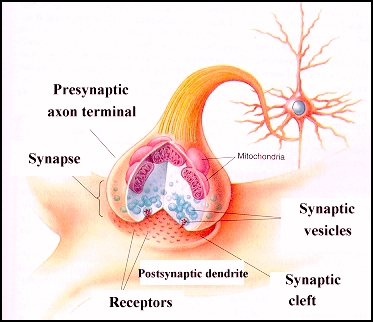
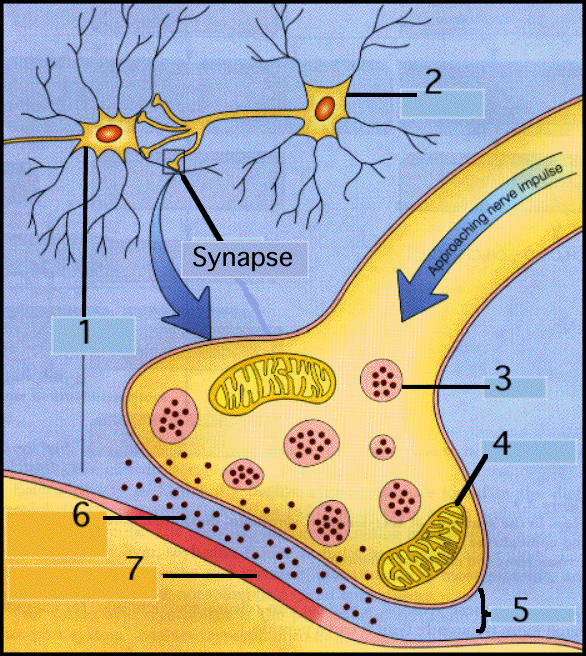
La sinapsis es un punto de machimbre o de enlace entre dos neuronas, la presináptica y la postsináptica. Las fibras nerviosas actúan como terminales de bujías eléctricas de los motores de explosión. Hay una luz o una brecha sináptica entre los terminales, brecha sináptica donde descargan vesículas sinápticas que difunden, ayudan a que ocurran reacciones físicas y químicas, recapturan los neurotransmisores ya usados y propagan potencial eléctrico desde una pared o membrana de la brecha o hendidura, la de la neurona presináptica, a la pared o membrana de la otra, la postsináptica.En su extremo, el axon de los nervios se ramifica en muchos terminales pequeños que llegan a estar en contacto estrecho con las dendritas de otras neuronas. Al contacto entre dos neuronas se le llama sinapsis. El axon y la dendrita nunca se tocan. Siempre hay un pequeño vacio llamado hendidura sinaptica. Cuando la señal electrica llega a un terminal nervioso, hace que el nervio libere neurotransmisores. Los neurotransmisores son agentes quimicos que viajan una corta distancia hasta las dendritas mas próximas.
A la neurona que libera el neurotransmisor se le llama neurona presinaptica. A la neurona receptora de la señal se le llama neurona postsinaptica. Dependiendo del tipo de neurotransmisor liberado, las neuronas postsinapticas son estimuladas (excitadas) o desestimuladas (inhibidas). Cada neurona se comunica con muchas otras al mismo tiempo. Puesto que una neurona puede enviar o no un estimulo, su comportamiento siempre se basa en el equilibrio de influencias que la excitan o la inhiben en un momento dado. Las neuronas son capaces de enviar estimulos varias veces por segundo.La sinapsis es un hecho comunicativo entre dos neuronas, una presináptica y otra postsináptica. Es imprescindible la conducción previa del impulso nervioso en la neurona presináptica y particularmente, en los denominados botones terminales, que son las últimas estructuras de la ramificación y diversificación axónica de la neurona presináptica. Esta circunstancia es el primer punto de acción para los fármacos y drogas que afectan a la sinapsis, pues en concreto, la modificación de la conductibilidad, aun no siendo un fenómeno tan asequible como otras etapas de la sinapsis, es uno de los caminos para la intervención de anestésicos que infiltrados a distintas concentraciones bloquean o modifican la conductibilildad.
Algunos neurotransmisores como acetilcolina (ACh), glicina, glutamato, aspartato y ácido gamma-amino butírico (GABA), tienen una actividad biológica directa aumentando la conductancia a ciertos iones por adherencia a canales iónicos activados en la membrana postsináptica. Otros neurotransmisores, como la noradrenalina (NA), dopamina (DA) y serotonina (5-HT), no tienen actividad directa pero actúan indirectamente vía sistemas de segundo mensajero para causar la respuesta postsináptica. Estos sistemas implican adenosín-monofosfato-cíclico (AMPc), guanidín-monofosfato-cíclico (GMPc), inositol trifosfato (ITP), diacil glicerol (DAG), prostaglandinas (Pgs), leucotrienos, epóxidos y Ca++.
4.2.3.1. Clases de sinapsis.
La función de la neurona es la comunicación y la función del SN es generar un comportamiento, ambos en virtud de las conexiones interneuronales. Una neurona ejerce su influencia para excitar a otras neuronas mediante los puntos de unión o sinapsis. Cada unión sináptica está formada por una parte de una neurona (terminal sináptico) que conduce un impulso a la sinapsis y por otra, de otra neurona (estructura postsináptica) que recibe el impulso en la sinapsis (Barr, 1994).El impulso nervioso debe atravesar un espacio muy pequeño (20nm), denominado hendidura sináptica que separa las estructuras pre y postsinápticas y puede propagarse en cualquier dirección por la superficie de la neurona; sin embargo la dirección que toma en condiciones fisiológicas se determina por una polarización constante que se hace en la sinapsis, donde la transmisión se efectúa del axón de una neurona a la superficie de otra neurona (Darnell, 1993).
La dinámica estructural y funcional para que se lleve a cabo una sinapsis entre dos neuronas esta dada por el movimiento, descarga, recaptación y reformación (resíntesis) de un neurotransmisor. Algunos neurotransmisores, como los péptidos, por ejemplo, son producidos en el soma, empaquetados en las vesículas que migran a través del axón mediante flujo axoplásmico por medio de los microtúbulos hacia el terminal presináptico donde se conocen con el nombre de vesículas sinápticas. La naturaleza del contenido de la vesícula sináptica varía dependiendo de la región cerebral de donde esta proceda. Cuando un impulso llega al terminal sináptico esta acompañado por la entrada de iones calcio en el citoplasma neuronal. El calcio proviene de los fluidos tisulares que están fuera de la neurona (espacio extracelular), estos iones, que han atravesado la membrana celular se unen a la molécula transportadora. Los iones calcio impulsan la migración de alguna de las vesículas sinápticas hacia la membrana presináptica, la membrana de cada vesícula sufre un proceso de fusión como la membrana presináptica, lo cual esta seguido por una expulsión rápida exocitosis del neurotransmisor libre en la hendidura sináptica (Meyer, 1985).
El neurotransmisor liberado en la hendidura sináptica interaccciona directamente con las moléculas del receptor en la membrana postsináptica. Mediante este tipo de interacción se abren un gran número de canales iónicos específicos que permiten el flujo de una corriente eléctrica, transportada por iones cargados a través de la membrana postsináptica lo que afecta al estado electroquímico de la membrana en el área inmediata al canal. De esta forma la excitabilidad eléctrica de esta pequeña porción de membrana puede aumentar o disminuir mediante despolarización o hiperpolarización de la misma. Las alteraciones eléctricas individuales de la membrana postsináptica ejercen un efecto en el potencial de membrana de la neurona, que puede llevar a la generación del impulso nervioso (Barr, 1994).
En algunos casos cuando se libera el neurotransmisor, este es inactivado generalmente por hidrólisis, los fragmentos que resultan del neurotransmisor se eliminan del sistema o se reciclan por endocitosis, en cuyo caso, los fragmentos se reincorporan a una nueva vesícula formada en la membrana presináptica. Este tipo de vesícula se conoce como vesícula encapsulada y tiene apariencia diferente a la vesícula sináptica. Los fragmentos que entran en la neurona por endocitosis son utilizados para la resíntesis del neurotransmisor (Cooper, 1994).
Existen algunos principios generales para la identificación de dos tipo de sinapsis: excitadoras e inhibidoras. Datos electrofisiológicos, muestran la distribución de las sinapsis excitadoras a nivel de la porción superior del árbol dendrítico de las neuronas centrales y de las sinapsis inhibidoras que habitualmente están unidas a los segmentos iniciales de las dendritas o a los cuerpos celulares (Bradford, 1988).
A partir de la excitabilidad de las neuronas, que es su propiedad específica, se desencadenan distintos tipos de mecanismos que trascienden a la propia neurona y que establecen una clara comunicación entre las mismas. Esto es a lo que denominamos sinapsis, una región celular clara, concreta y muy estructurada definida por el mantenimiento de un espacio interneural, y cuyo significado final es el de la comunicación interneural a la que nos referimos en términos generales como sinapsis eléctrica y sinapsis química. En el primer caso, se habla siempre de una comunicación excitatoria con la continuidad de la conducción de la excitabilidad o el impulso nervioso; en el segundo caso se habla de una comunicación excitatoria o inhibitoria mediada por una sustancia química que no es otra cosa que el neurotransmisor. La sinapsis es, sin lugar a dudas, la estructura más lábil y accesible a las distintas sustancias químicas como fármacos y drogas y, por tanto, el lugar y término de referencia más importante en los mecanismos de acción de los distintos psicofármacos.
La sumatoria de los impulsos exitatorios e inhibitorios que llegan por todas las sinapsis que se relacionan con a cada neurona( 1000 a 200.000) determina si se produce o no la descarga del potencial de acción por el axón de esa neurona
Las sinapsis suelen clasificarse en dos tipos según la transmisión del impulso: sinapsis eléctrica y sinapsis química.4.2.3.2. Sinapsis eléctrica.
En este tipo de sinapsis los procesos pre y postsináptico son continuos (2 nm entre ellos) debido a la unión citoplasmática por moléculas de proteínas tubulares a través de las cuales transita libremente el agua, pequeños iones y moléculas por esto el estímulo es capaz de pasar directamente de una célula a la siguiente sin necesidad de mediación química (Barr, 1994). Corresponden a uniones de comunicación entre las membranas plasmáticas de los terminales presináptico y postsinápticos, las que al adoptar la configuración abierta permiten el libre flujo de iones desde el citoplasma del terminal presinático hacia el citoplasma del terminal postsináptico.
La sinapsis eléctrica ofrece una vía de baja resistencia entre neuronas, y hay un retraso mínimo en la transmisión sináptica porque no existe un mediador químico. En este tipo de sinapsis no hay despolarización y la dirección de la transmisión está determinada por la fluctuación de los potenciales de membrana de las células interconectadas (Bradford, 1988).4.2.3.3. Sinapsis química.
La mayoría de las sinapsis son de tipo químico, en las cuales una sustancia, el neurotransmisor hace de puente entre las dos neuronas, se difunde a través del estrecho espacio y se adhiere a los receptores, que son moléculas especiales de proteínas que se encuentran en la membrana postsináptica (Bradford, 1988).
La energía requerida para la liberación de un neurotransmisor se genera en la mitocondria del terminal presináptico. La unión de neurotransmisores a receptores de la membrana postsinápticas produce cambios en la permeabilidad de la membrana. La naturaleza del neurotransmisor y la molécula del receptor determina si el efecto producido será de excitación o inhibición de la neurona postsináptica (Barr, 1994). Se han descrito varias formas de sinapsis según las estructuras implicadas. (Bradford, 1988).Axosomática: Sinapsis entre un axón y un soma.
Axodendrítica: Sinapsis ocurrida entre un axón y una dendrita.
Axoespinodendrítica: Sinapsis entre un axón y una espina dendrítica.
Axoaxónica: Sinapsis entre dos axones.
Dendrodendrítica: Sinapsis ocurrida entre dos dendritas.
Somatosomática: Sinapsis entre dos somas.
Dendrosomática: Sinapsis entre un soma y una dendrita.Existen dos clases de sinapsis química: la sinapsis asimétrica o tipo I se caracteriza por la diferencia en densidad de las membranas presináptica y postsináptica, siendo más gruesa la última. Esta densidad consiste de un material proteico que puede estar asociado al receptor postsináptico; la sinapsis simétrica o tipo II se caracteriza porque las membranas presináptica y postsináptica poseen un grosor semejante (Bradford, 1988).
4.2.3.4. Mecanismos de exitación e inhibición sinaptica.
El líquido extracelular tiene una gran cantidad de concentración de iones de sodio en cambio de la concentración de iones potasio es baja, por otra parte en el citoplasma de la neurona hay una alta concentración de iones potasio y una baja concentración de iones sodio. En estado de reposo los iones sodio pueden salir de la célula por difusión mediante los canales de la membrana. Unicamente pequeñas cantidades de sodio se difunden a través de la membrana, la cual, en reposo, es poco permeable a estos iones, grandes cantidades de iones sodio penetran en el momento de la conducción de los impulsos la entrada de sodio y la pérdida de potasio intracelular encuentran la oposición de distintas proteínas de la membrana constituyendo la bomba de sodio-potasio. La bomba la produce la molécula rica en energía, el ATP, para transportar iones a través de la membrana contra un gradiente de concentración, así las concentraciones de iones en el citoplasma se mantienen mediante gasto de energía que se producen de manera importante por consecuencia de la actividad de la bomba. La diferencia que resulta de las concentraciones de iones producen en la membrana un potencial de reposo, con el interior de la neurona que alcanza aproximadamente menos 70 mV con respecto al exterior (Zimmermann, 1993).
Durante la excitación que se atribuye a una variedad de estímulos ya sean químicos o físicos, se presenta una reducción del potencial de membrana y se dice que dicha membrana se despolariza. La reducción del potencial se extiende en dirección lateral en la membrana disminuyendo su magnitud con la mayor distancia a partir del punto de iniciación. Este cambio en el grado de potencial, es el tipo único de señalización en las dendritas y en el cuerpo de la neurona (Darnell, 1993).
La producción de estímulos en cantidad e intensidad suficiente puede reducir el potencial de membrana en el segmento inicial del axón hasta 10 a 15 mV. Este es un valor que desencadena la abertura de los canales de sodio regulado por el voltaje de la membrana del axón, los iones aparecen localmente en la superficie exterior procedentes de la superficie interna disminuyendo el gradiente y la concentración y atraídos por el exceso de carga negativa en el axoplasma. El interior del axón es temporalmente de +40 mV respecto al exterior. Este cambio se llama el potencial de acción. Una vez que se generó este potencial se propaga por la membrana, en circuitos locales de corriente eléctrica, que abren los canales cercanos de sodio. El potencial de acción al desplazarse produce un impulso nervioso (Meyer, 1985).
Se dice que los estímulos que despolarizan la membrana de la neurona son excitatorios, debido a que cantidad suficiente de ellos iniciarán el potencial de acción. Algunos estímulos originan el efecto opuesto de hiperpolarización, en este caso el potencial de membrana excede la cifra del potencial de reposo -70mV. Los impulsos que causan hiperpolarización inhiben la generación de potenciales de acción, debido a que se oponen a los efectos de los estímulos despolarizantes (Zimmermann, 1993).Primera etapa: Regulación de la síntesis del neurotransmisor.
Un botón terminal en equilibrio mantiene una polarización de su membrana y una regulación de su capacidad de síntesis del neurotransmisor o de las proteínas asociadas a la consolidación de las vesículas sinápticas, de acuerdo con las exigencias funcionales de la neurona y de su propio control metabólico sobre la disponibilidad de neurotransmisor.
El neurotransmisor puede almacenarse en vesículas sinápticas, pequeños reservorios globulares que contienen receptores en su pared exterior y permiten que el neurotransmisor se libere en sitios específicos de la terminal presináptica. Se ha hecho la analogía de la terminal presináptica como un espacio donde las vesículas sinápticas, así como las mitocondrias y otras estructuras subcelulares, están flotando. Sólo en un lugar determinado de este espacio se localizan los sitios por donde el neurotransmisor puede liberarse hacia el exterior. Como si las vesículas fueran huevos que sólo pudieran acomodarse en los huecos de sus cajas, que se encuentran en el piso de este espacio. Y sólo a través de los huecos de estas cajas se puede descargar el contenido hacia el exterior. En este caso, los huecos tienen receptores que reconocen los componentes de la cáscara del huevo.
La presencia de estas vesículas y de receptores en sus membranas, y de moléculas de el neurotransmisor significa que existe todo lo necesario en el interior de la terminal para sintetizar todos estos componentes, y de mecanismos para la regulación de esta síntesis y de la liberación sináptica. La terminal presináptica tiene autorreceptores que le informan sobre los niveles del neurotransmisor en el exterior de la terminal. Si sus niveles son elevados, la terminal puede fabricar o liberar menos. Si éstos son bajos, puede hacer lo contrario. La activación, pues, de los autorreceptores puede tener efectos de estimulación o, generalmente, de inhibición de la liberación sináptica. La membrana de la terminal presináptica también posee moléculas transportadoras del mismo neurotransmisor que ellas liberan. Es un mecanismo de recaptación que sirve no sólo para ahorrar neurotransmisor al reutilizarlo, sino también contribuye a limitar sus efectos postsinápticos.
Asumiendo esta condición, la conducción de un impulso nervioso, que rompe el esfuerzo fisiológico para el mantenimiento de la polarización, produce una significativa movilización de las vesículas sinápticas que almacenan neurotransmisores, los cuales llegan a unirse a la propia membrana plasmática; y la entrada de Ca++, como consecuencia de la despolarización, que abre los canales de Ca++, rompe la anastomosis vesícula-membrana y libera al espacio sináptico el neurotransmisor. El aumento de la superficie de membrana que supone la ruptura de las vesículas sinápticas, es posteriormente restaurado por la pinocitosis interna, es decir, el desprendimiento interno de estas membranas que van a ser, desde luego, precursoras para la formación de nuevas vesículas sinápticas. Asimismo, el Ca++ que ha intervenido en el proceso es transportado extracelularmente por las proteínas transportadoras de Ca++ similares a la bomba de sodio-potasio, con lo que se vuelve al equilibrio en la neurona presináptica.Segunda etapa: Liberación del neurotransmisor.
Una vez que el neurotransmisor ha sido liberado al espacio sináptico, se difunde por el mismo y puede seguir las siguientes rutas:
Tercera etapa: Activación de receptores.
- Fijación en los lugares específicos de membrana tanto presináptica como postsináptica, que son los receptores.
- Dispersión en el espacio sináptico y actuación fuera de la sinapsis como un neuromodulador.
- Recaptación presináptica del neurotransmisor.
- Catabolización enzimática del neurotransmisor y por tanto degradación de la estructura.
Receptores postsinápticos
Las estructuras de unión del neurotransmisor en la membrana postsináptica son proteínas específicas de membrana que constituyen los receptores postsinápticos y que son la clave del reconocimiento de la liberación de neurotransmisores presinápticos. Es decir, los receptores postsinápticos son la estructura básica para interpretar la comunicación interneuronal, constituyen, por tanto, auténticas estructuras de diferenciación neuronal a la vez que son un claro exponente de la diversidad específica.
La afinidad del neurotransmisor como ligando por el receptor postsináptico es una característica inherente a la diferenciación y especialización de las propias neuronas, hasta tal punto que un neurotransmisor determinado no produce efectos, por muy constante e intensa que sea su liberación, si la membrana postsináptica no contiene en su diferenciación los receptores específicos para ese neurotransmisor concreto.
Una vez que el receptor potsináptico y neurotransmisor se unen, aparece inmediatamente una consecuencia en la membrana postsináptica. Indudablemente, hay una alteración constitucional que puede ser suficiente para modificar los canales iónicos de polarización de esa membrana, generando rápidamente una despolarización o una hiperpolarización, lo que conllevaría un aumento de la activación o inhibición, respectivamente, de la membrana postsináptica.
Normalmente, la activación de los receptores postsinápticos por un neurotransmisor ejerce su efecto, es decir, es interpretada postsinápticamente, mediante la utilización de un neuromediador o de un segundo mensajero. En el primer caso, lo que ocurre es que la acción neurotransmisor-receptor es indirecta y necesita de una proteína de membrana mediadora ante la modificación de su permeabilidad iónica, o lo que es lo mismo, hacia la alteración de su polarización. En el segundo caso, bien por la neuromediación, bien por el complejo específico neurotransmisor-receptor, se activa un sistema enzimático de membrana, normalmente la adenilato-ciclasa, cuya actuación inmediata se realiza sobre el adenosín trifosfato (ATP) que se transforma en adenosín monofosfato cíclico (AMPc). El AMPc no sólo activa la apertura de los canales iónicos sino que también conduce la información de la actividad de membrana de la que procede, hacia las estructuras somáticas y nucleares de la neurona, por lo que las consecuencias de una comunicación interneuronal pueden perfectamente trascender a mecanismos bioquímicos que impliquen cambios estructurales y/o funcionales a partir de la utilización del segundo mensajero.
Al igual que ocurría en la segunda etapa, es decir, la dependencia del Ca++ en la liberación del neurotransmisor, todavía aquí nos encontramos con una incuestionable dependencia del Ca++ que es tanto más acusada cuanto mayor es la repercusión metabólica del AMPc.
Como resultado final de esta etapa, lo que ocurre localmente es una movilización de los canales iónicos que afectan al Na+, K+, Cl- y Ca++. Como sabemos, la polarización de la membrana establece particularmente unas concentraciones de Na+ extracelular y de K+ intracelular. Si se abre el canal de Na+, aparece una única entrada que genera automáticamente una despolarización, es decir crea un potencial postsináptico excitador. Si lo que se activa es el canal de K+, se establece una acumulación excesiva de K+ intracelular cuya consecuencia es la contraria, es decir, una hiperpolarización de la membrana y por tanto un potencial postsináptico inhibidor. La acción sobre el canal de Cl- tiene efecto cuando la membrana está ligeramente despolarizada, en cuyo caso la apertura de este canal se opone a la aparición de un potencial postsináptico excitador. Cuando la membrana está polarizada, cuyo resultado final es una clara electronegatividad intracelular, la apertura de un canal de Cl- no tiene mayor repercusión al reafirmar la electronegatividad, y además es rápidamente reequilibrada por difusión.
Por su parte los iones Ca++ situados extracelularmente actúan igual que los iones Na+; es decir, despolarizan la membrana y producen potenciales postsinápticos excitatorios, además de ser un potente activador enzimático.Receptores presinápticos o autorreceptores
Es también enormemente transcendente la activación de los receptores específicos de membrana, situados en la neurona presináptica, que responden a la liberación del neurotransmisor con la misma eficacia y afinidad que los receptores postsinápticos. Ahora bien, el resultado de este mecanismo es un servomecanismo, es decir, la autorregulación de la liberación del neurotransmisor con el fin de que la comunicación neuronal sea eficaz, ajustada y esté adecuada a los niveles de excitabilidad que han provocado los potenciales de acción en los botones terminales. Estos receptores, a diferencia de los postsinápticos, no controlan canales iónicos. Sin embargo, sí que regulan la movilización de las vesículas, intervienen esencialmente en la disponibilidad del neurotransmisor y, por tanto, en su síntesis y en su liberación. Normalmente, su control es por retroalimentación y su activación favorece los mecanismos de inhibición presináptica que, lógicamente, se asientan en el funcionamiento de los segundos mensajeros.Cuarta etapa: Inactivación del neurotransmisor
Una vez cumplido el objetivo sináptico, es decir la comunicación a la neurona postsináptica, se produce la inactivación del neurotransmisor. El mecanismo de inactivación más importante es la recaptación presináptica, que se produce no por la utilización de receptores específicos para su fijación, sino por auténticos transportadores de membrana que vuelven a incorporarlos al espacio presináptico. Estos transportadores de membrana actúan bien sobre el neurotransmisor íntegramente liberado y reconocido por sus receptores o bien sobre metabolitos intermediarios fruto de la acción catabólica de sistemas enzimáticos postsinápticos. Indudablemente, la recaptación o la degradación catabólica del neurotransmisor termina con la activación de los receptores de membrana y desde luego termina con la actividad de los neurotransmisores.
Quinta etapa: Difusión del neurotransmisor.
La ultima etapa que podemos definir en la sinapsis es la difusión del neurotransmisor. Esto nos permite incluir el concepto de neuromodulador, que produce sus efectos a grandes distancias del lugar de secreción, y que afecta a grupos neuronales y otras áreas de intervención. Este concepto es fundamental en la secreción de neuropéptidos más que de neurotransmisores clásicos, y aunque los propios neuropéptidos de secreción sináptica pueden actuar como auténticos neurotransmisores, la propia estructura de la molécula favorece su difusión y alejamiento de su actuación local, por lo que pueden ser considerados como neuromoduladores.
La naturaleza de la materia transmisora no es en realidad quien define el efecto postsináptico de la neurotransmisión, sino que es la naturaleza de los canales iónicos controlados por los receptores de membrana postsinápticos la que define el efecto excitatorio o inhibitorio; de ahí que podamos encontrarnos con efectos claramente inhibitorios de un determinado neurotransmisor que en su generalidad es excitatorio.
ttp://www.biopsicologia.net/fichas/page_8.html
http://omega.ilce.edu.mx:3000/sites/ciencia/volumen3/ciencia3/130/htm/sec_9.htm